|
Sind Vogelflügel umgestaltete Dinosaurierhände?
Zum Konflikt zwischen fossilen und entwicklungsbiologischen Daten
bei der phylogenetischen Herleitung eines Vogelflügels von Henrik Ullrich
Studium Integrale Journal
15. Jahrgang / Heft 1 - April 2008
Seite 18 - 30
|
 |
Zusammenfassung: Die phylogenetische Ableitung der Vögel aus einer Gruppe der Theropoden-Dinosaurier zählt zu einem häufig zitierten und wichtigen Baustein in evolutionären Modellvorstellungen. Betrachtet man aber im Detail die dafür relevanten Befunde, trübt sich das scheinbar klare Bild schnell ein, denn die Befunde erlauben eine vielfältige und kontroverse Deutung. In diesem Artikel werden relevante Daten aus der Embryologie zur Gliedmaßenentwicklung bei Wirbeltieren vorgestellt. Deren Bedeutung für Homologiebetrachtungen und für kausale Erklärungsansätze im Rahmen moderner Evo-Devo Konzepte werden diskutiert. Die große Spannbreite der diesbezüglich in der Literatur äußerst kontrovers vertretenen hypothetischen Vorstellungen führt beim gegenwärtigen Stand des Wissens zu folgenden Schlussfolgerungen:
- Es fehlt ein methodisch sicheres Kriterium, um aus mehreren möglichen Homologiezuweisungen auf der Basis embryologischer und paläontologischer Daten die phylogenetisch relevante zu extrahieren. Keine der vorgestellten Ansätze kann deshalb für sich ein höheres Maß an Plausibilität beanspruchen als andere. Die Homologiebeziehungen zwischen dem Vögelflügel und der vorderen Extremität von Dinosauriern bleiben unter phylogenetischer Perspektive auch nach 150 Jahren Evolutionsforschung unklar.
- Die kausalen Erklärungsversuche zur Entstehung des Grundbauplans des Vogelflügels auf der Grundlage der Synthetischen Evolutionstheorie sowie unter Rückgriff auf neue Ansätze der evolutionären Entwicklungsbiologie (Evo-Devo) sind als spekulative Extrapolationen von Details des verfügbaren embryologischen bzw. molekulargenetischen Wissens zu werten.
- Alle vorgelegten Hypothesen zur Evolution des Vogelflügels sehen sich drei defizitären Situationen ausgesetzt: Fehlendes Wissen über Wirkmechanismen bzw. die Wechselbeziehungen von äußeren und inneren Selektionsdrücken auf den evolutionären Umbau der vorderen Extremität und die vielfach noch unverstandenen genetischen und epigenetischen Regulationskaskaden der Gliedmaßenentwicklung während der Individualentwicklung bei Wirbeltieren. Drittens muss offen gelassen werden, welche Kausalitäten den phylogenetischen Wandel ontogenetischer Entwicklungsprogramme bedingen und ermöglichen.
- Die wissenschaftliche Klärung der homologen Beziehungen zwischen den Fingerstrahlen an Vogelflügeln und denen bei anderen Landlebewesen (insbesondere Echsen und Säugetieren) und ihre kausale Begründung bleibt eine der größten Herausforderungen für die Evolutionsforschung.
- Das ontogenetische und phylogenetische Modellobjekt „Flügelentwicklung“ ist exemplarisch für grundsätzliche Probleme und Grenzen der evolutionsgeleiteten Hypothesenbildung sowie darauf aufbauender Homologiebetrachtungen.
| |
|
1. Einführung
Die Analyse der ontogenetischen Entwicklung von Gliedmaßen bei Wirbeltieren zählt gegenwärtig zu einem der wichtigsten und erfolgreichsten Forschungsschwerpunkte der Embryologie. Der spezifisch-komplexe und gleichzeitig modulare Aufbau (Anatomie) von Flossen, Armen, Händen, Beinen, Füßen oder Flügeln gilt als exzellentes Modell, um fundamentale entwicklungsbiologische Fragestellungen zu klären. Zu diesen zählen beispielsweise die molekularbiologischen Grundlagen der Festlegung (= Speicherung) und der Realisierung (= Abrufen und Umsetzen) des Körperbauplans eines Organs während der Embryonalentwicklung oder die Steuerung der präzisen Anordnung von spezialisierten Zellen und der Gewebe in ihnen (Tickle 2006).
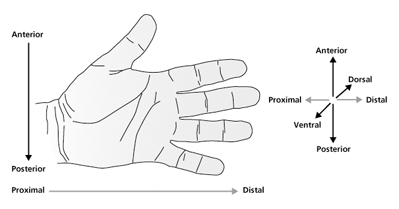 |
| Abb. 1: Die drei anatomischen Achsen zur Beschreibung der Lageverhältnisse von Strukturen der menschlichen Hand. Genau abgestimmte Entwicklungsprozesse der Embryonalentwicklung garantieren, dass der Daumen bzw. der kleine Finger immer am richtigen Ort entstehen. (Nach Tickle 2006) |
|
Das Extremitätenskelett der Wirbeltiere wird durch eine definierte Zahl von Knochen mit einer jeweils charakteristischen Größe und Form in einer festen räumlichen Anordnung gekennzeichnet. Anhand der drei orthogonalen Raumachsen (Höhe, Breite, Tiefe) lassen sich die einzelnen Bestandteile der Gliedmaßen zueinander während der embryonalen Bildung und im ausgewachsenen Zustand beschreiben. Zum besseren Verständnis der weiteren Ausführungen sei dies am Beispiel der Hand des Menschen kurz illustriert (Abb. 1). Die proximo-distale Achse verläuft von der Schulter zu den Fingerspitzen, also von körpernah zu körperfern, die anterio-posteriore (vorn-hinten) Achse vom Daumen (Finger I) zum kleinen Finger (Finger V) und die dorso-ventrale (Rücken-Bauch) Achse vom Handrücken zur Handfläche. Die zahlreichen genetischen und epigenetischen Prozesse, welche die Ontogenese der Extremitäten in allen drei Raumachsen regulieren, verlaufen hochgradig koordiniert, vernetzt und einander überlagernd. Zahlreiche Details zur Differenzierung und Strukturbildung wurden in den letzten Jahrzehnten erhellt (z.B. Herkunft der einzelnen Zelltypen wie Muskelzellen, Nervenzellen, Blutgefäßzellen), einige werden kontrovers diskutiert (z.B. die Grundlagen der proximo-distalen Differenzierung) und etliche Prozesse der Musterbildung liegen noch im Dunkeln (z.B. Spezifikation von Form und Anzahl der Fingerstrahlen).
Die Ontogenese der Wirbeltierextremitäten gewinnt zunehmend auch hinsichtlich phylogenetischer Fragestellungen an Bedeutung. Im Blickpunkt stehen dabei vergleichende Untersuchungen der Fingerentwicklung zur Feststellung von Homologien, die der Stammbaumrekonstruktion – insbesondere zur Klärung der phylogenetischen Ableitung der Vögel – dienen. Darüber hinaus scheint das ontogenetische Modellobjekt „Extremitätenentwicklung“ gut dafür geeignet, die deskriptiv abgeleiteten überartlichen Homologien kausal zu erklären, z.B. durch evolutionäre Änderungen embryonaler Entwicklungsprozesse auf dem Weg homeotischer Mutationen. Dieser Ansatz wird gegenwärtig durch die Evolutionsforschung favorisiert, um den postulierten Wandel des Bauplanes der vorderen Extremität einer landlebenden Echse (z.B. Theropode) in den Flügel eines Vogels verständlich zu machen.
Der vorliegende Artikel möchte aus embryologischer Perspektive einen Überblick zum Stand der Diskussion hinsichtlich der Ableitung der Vögel von den Dinosauriern geben. Dazu werden zunächst kurz die etablierten und konkurrierenden paläontologischen Sichtweisen vorgestellt. Eine Einführung in die wesentlichen Vorgänge, die zur Musterbildung und zur Differenzierung von Strukturen der vorderen Extremität bei Vögeln und Säugetieren während der ontogenetischen Entwicklung führen, ist Voraussetzung, um verschiedene phylogenetische Konzeptionen der Flügelentstehung zu erklären und zu bewerten. Abschließend soll begründet werden, warum die Integration ontogenetischen Wissens inhaltlich und methodisch nur wenig dazu beitragen konnte, die vielen offenen Fragen und widersprüchlichen Interpretationen bei der Ableitung der Vögel von den Dinosauriern aufzulösen.
|
Amnioten: Landwirbeltiere, deren Embryonen sich in einer mit Fruchtwasser (Amnionflüssigkeit) gefüllten Amnionhöhle entwickeln, die von einer Embryonalhülle (Amnion) umgeben ist (Reptilien, Vögel, Säugetiere). Archosaurier: Als Archosauria („Herrscherreptilien“) wird eine Reihe von ausgestorbenen Reptiliengruppen (u.a. Dinosaurier) sowie die heute noch lebenden Krokodile (Crocodylia) und Vögel (Aves) zusammengefasst. Cladistik, cladistisch: auf quantifizierbare und numerische Daten basierende phylogenetische Taxonomie, in der alle verfügbaren Merkmale von Organismen gleichberechtigt berücksichtigt werden, um Verwandtschaftsbeziehungen („ursprünglich“ und „abgeleitet“) darzustellen. Für die Artbildung wird ausschließlich eine dichotome Verzweigung der Stammart zugelassen. deskriptiv: beschreibend. Determinierung: In der Entwicklungsbiologie Beschreibung der irreversiblen Festlegung von entwicklungsfähigen Zellen auf einen spezifischen Endzustand (Muskelzelle, Knochenzelle). Ektoderm: äußere Zellschichten des embryonalen Keims bei Wirbeltieren, aus dem später u.a. Haut und Anteile des Nervensystems hervorgehen. Embryologie: Biologische Wissenschaft, welche die individuellen Entwicklungsvorgänge des mehrzelligen Organismus beschreibt und kausal zu erklären versucht. Epigenetische Faktoren: in Ergänzung zum genetisch vorgegebenen Informationspool alle anderen informationstragenden und informationsverarbeitenden zellulären Strukturen (z.B. Proteine, Zellorganellen, Zellmembranen). Erweiterte Synthetische Evolutionstheorie: Nach Kutschera (2006) Verbindung der Synthetischen Evolutionstheorie u.a. mit den aktuellen Ergebnissen der Geologie, Molekularbiologie, Computersimulation und Entwicklungsbiologie. Evo-Devo: Moderner Forschungszweig der Evolutionsbiologie, besonderer Schwerpunkt liegt in der Integration aus der Embryologie stammender Erkenntnisse und ihrer überartlichen Bewertung bei der Beschreibung und Erklärung evolutionärer Prozesse. FGF: fibroplast-growth-Factor, besondere Gruppe regulatorisch wirksamer Eiweiße während der Ontogenese (Morphogene). Genetische Expressionsmuster: räumlich und zeitliches fassbares Muster von Aktivitäten einzelner Gene in einer morphologischen Struktur. Genotyp: Gesamtheit der genetischen Erbfaktoren. Homeotische Gene: Gene, die in der Ontogenese u.a. an der Koordination des Erscheinungsortes von morphologischen Merkmalen (z.B. Armanlage, Antenne bei Drosophila) oder bei seriell auftretenden Strukturen (Wirbelkörper, Pharyngula) mitwirken. Homologie: Gleichwertigkeit ähnlicher Strukturen, Organe, Gene und Prozesse; in der Evolutionsbiologie Beleg für gemeinsame Abstammung (Extremitäten der Landwirbeltiere). homöotische Mutationen: Mutationen, die während der Ontogenese dazu führen, dass morphologische Merkmale an einem „falschen“ Ort (z.B. Antenne bei Drosophila am Platz des Beines) oder seriell auftretende Strukturen sich zahlenmäßig und/oder in ihrem Charakter verändern (z.B. Umwandlung von Brustwirbelkörper in einen Lendenwirbelkörper). Hox-Gene: In der Ontogenese wichtige Gruppe regulatorisch wirksamer Gene, enthalten einen identischen Genabschnitt, der für einen spezifischen Proteinabschnitt (Homeobox) codiert. Konvergenz: Funktionelle oder anatomische Ähnlichkeit von Strukturen, Organen, Genen und Prozessen stammesgeschichtlich nicht verwandter Organismen; von der Evolutionsbiologie als Parallelentwicklungen interpretiert (z.B. Körperform von Pinguin, Hai, Fischsaurier, Delphin). Masterkontrollgene: Gesamtheit aller Gene, denen in der Ontogenese eine wichtige regulatorische Funktion zugeschrieben wird (z.B. Hox-Gene, PAX-Gene). Mesenchym: Aus dem mittleren Keimblatt des embryonalen Keims bei Wirbeltieren hervorgehende Strukturen, wie Gefäße, Knochen, Bindegewebe. Morphogene: Besondere Gruppe von Eiweißen, die als Transkriptionsfaktoren oder regulativ wirksame Proteine die Expression von Strukturgenen beeinflussen. Morphologie: Lehre vom Aufbau und der Gestalt der Lebewesen. Ontogenese: Individualentwicklung eines Organismus, bei Wirbeltieren von der Befruchtung bis zur Geburt/Schlüpfen des Jungtieres dauernder Zeitabschnitt. Phänotyp: Gesamtheit der äußerlichen Gestaltmerkmale eines Organismus. Phylogenese: Stammesgeschichte. prächondrale Anlagen: der Knorpelbildung vorangehende Struktureinheiten aus entsprechenden bindegewebigen Komponenten. rudimentär: in der Evolutionsbiologie genutzt um phylogenetisch rückgebildete Organe zu kennzeichnen (z.B. Wurmfortsatz); hier: Anlage einer ontogenetischen Struktur, die bei anderen Individuen eine weitere Entwicklung erfahren. Somiten: segmental angeordnete Bindegewebsblöcke, aus denen während der Ontogenese unter anderem die Wirbelkörper hervorgehen. Theropoden-Dinosaurier: Dinosauriergruppe (Saurischa) mit drei Fingerstrahlen, gelten als Vorfahren der Vögel. Transkriptionsfaktor: Proteine, die die DNS dazu anregen, sich identisch zu duplizieren bzw. ihre Information auf die mRNS zu übertragen.
|
|
|
2. Ableitung der Vögel von den Dinosauriern – Paläontologische Sichtweisen
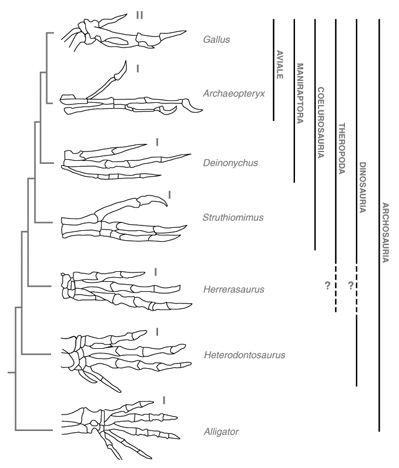 |
| Abb. 2: Schematische Illustration der Handelemente von verschiedenen Vertretern der Archosaurier. Die hier wiedergegebene phylogenetische Systematik orientiert sich an Gauthier (1986). Der vordere (anteriore) Finger ist entsprechend dem gegenwärtigen Konsens mit I oder II bezeichnet. Die jeweilige systematische Zuordnung wird rechts angegeben. Bestehende Unsicherheiten der systematischen Zuordnung werden durch ein „?“ gegekennzeichnet. (Nach Welten 2005) |
|
Überblick. Ausgedehnte phylogenetische Analysen fossiler Arten führten zu dem Schluss, dass Vögel als lebende theropode Dinosaurier anzusehen sind (Vargas & Fallon 2005). Die fünffingrigen Vorfahren der Theropoden verloren den vierten und fünften Strahl, was einige Funde mit reduzierten Anlagen des vierten und fünften Fingers belegen sollen. Außerdem besteht ein hoher Grad an Ähnlichkeit zwischen dem Bau der drei Fingerstrahlen des „Urvogels“ Archaeopteryx und dem von Theropoden. Aus vergleichender paläontologischer Sicht werden deshalb die drei Strahlen des Vogelflügels als homolog zu den ersten drei Fingerstrahlen theropoder Dinosaurier bewertet. Diese Interpretation wird allerdings nicht von allen Biologen geteilt (z.B. Feduccia et al. 2005).
Die Mehrheit der Forscher favorisiert auf der Basis cladistischer Analysen die Hypothese, dass die Vögel sich aus einer Untergruppe der Theropoden-Dinosaurier phylogenetisch herleiten. Zur wichtigsten Stütze für diese Vorstellung zählt die Ähnlichkeit der drei bekrallten Finger des Flügels von Archaeopteryx im Vergleich mit der Anatomie bei einigen dreifingrigen Theropoden, die eine Reduktion des IV. und V. Fingers zeigen (z.B. bei Deinonychus, Struthiomimus, Herrerasaurus).
Als Ausdruck der vermuteten homologen Identität zu den Theropoden bezeichnete man die Finger bei Archaeopteryx mit der römischen Nomenklatur I, II, III (I entspricht dabei dem homologen „Daumen“ der Krokodile oder Säugetiere). Diese Nomenklatur konnte aber nicht pauschal auf die Fingerstrahlen des Flügels bei modernen Vögeln übertragen werden, da der embryonale Befund, wie später erläutert wird, hier eine andere Kennzeichnung verlangt, nämlich mit II, III, IV (vgl. dazu Abb. 2).
Neuere Funde von Theropoden (speziell aus der Gruppe der Maniraptoren) mit Federn bzw. mit einer federähnlichen Bedeckung (z.B. Protoarchaeopteryx und Caudipteryx) wurden von zahlreichen Forschern als Bestätigung der Theropodenhypothese gewertet (z.B. Zhou et al. 2003). Diese Interpretationen sind jedoch sehr umstritten. Nach Peters (2002) und Paul (2002) sprechen die Befundkonstellationen bei den o.g. Formen eher dafür, dass es sich um sekundär flugunfähig gewordene Vögel handelt und sie nicht zu den Dinosauriern gehören. Feduccia et. al. (2005) konnten zeigen, dass die Interpretation von fossilen Resten der Hautbedeckung – wie z. B. bei dem Theropoden Sinosauropteryx – als „Protofedern“ nicht zwingend ist. Darüber hinaus kann die Theropodenhypothese bisher zahlreiche fundamentale Fragen der Vogelevolution nicht beantworten (Überblick bei Junker 2005). Auf der Grundlage paläontologischer Daten wurden deshalb auch als mögliche alternative Ausgangsformen unspezialisierte Thecodontier (eine Stammform der Archosaurier) und urtümliche krokodilähnliche Formen (Crocodilia) diskutiert (Hecht 1985; Moinar 1985; Walker 1985). Welten et al. (2005) verweisen zusätzlich auf die Möglichkeit, dass die Vögel nicht zwingend von einer pentadaktylen Urform (Formen mit fünf Fingerstrahlen) abgeleitet werden müssen. Polydaktyle Archosaurier, also Urechsen mit mehr als fünf Fingerstrahlen, sind theoretisch ebenso wahrscheinlich und wurden schon von Bardeleben (1889) und Holmgreen (1955) als Vorfahren der Vögel ins Gespräch gebracht. Diese Position besitzt aber gegenwärtig nur einen Außenseiterstatus.
|
|
3. Ontogenetische Prinzipien der Gliedmaßenentwicklung bei Vögeln und Landwirbeltieren
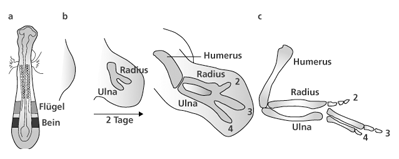 |
| Abb. 3: Die Entwicklung der Flügelknospe beim Hühnchen. a Anlage der Extremitätenknospen (Buds) in enger räumlicher Beziehung zu bestimmten Somiten, b Auswachsen der Flügelknospe in proximo-distaler Richtung, phasenweise Ausbildung der knorpligen Anlagen für die Oberarm-, Unterarm- und Handstrukturen, c Schematische Darstellung des knöchernen Grundbauplanes der oberen Extremität. (Nach Tickle 2006) |
|
Überblick. Das Studium der ontogenetischen Entwicklung der Extremitäten gilt als Modellobjekt für die Erforschung der Interaktionen zwischen Genotyp und Phänotyp bei der Etablierung einer komplizierten anatomischen Struktur unter dem Einfluss hochgradig vernetzter und raum-zeitlich agierender Steuerungssysteme. Die theoretischen und technischen Fortschritte der experimentellen Entwicklungsbiologie gestalteten die Forschungen in den letzten Jahren sehr erfolgreich. Auf der Basis des jeweils aktuellen Wissens wurden unterschiedliche Modelle („progress zone model“, „early specification model“) der ontogenetischen Gliedmaßenentwicklung entworfen. Die zunächst als sehr verheißungsvoll betrachtete Bedeutung der Hox-Gene hat sich relativiert. Diese sind zwar ein wichtiger Bestandteil eines mehrdimensional vernetzten Entwicklungssystems, aber nur eines unter vielen. Der stetige Wandel der örtlichen Expressionsdomänen von Hox-Genen in der Extremitätenknospe erlaubt keine feste Zuordnung einzelner Hox-Gene zu bestimmten morphologischen Strukturen. Das gegenwärtige Verständnis über die kausalen Zusammenhänge und Wechselbeziehungen genetischer bzw. epigenetischer Einzelprozesse, die zur Ausbildung der Gliedmaßen während der Ontogenese führen, bleibt weiterhin sehr lückenhaft. Die gewonnenen Erkenntnisse dienen als Grundlage für verschiedene phylogenetische Deutungen im Rahmen von Evo-Devo Konzeptionen.
Richardson et al. (2003) kennzeichneten die Individualentwicklung als einen vierdimensionalen Prozess. Drei Dimensionen betreffen die Veränderungen der Einzelmerkmale im Raum (Länge, Breite, Tiefe) und die vierte ihren Wandel in der Zeit. Der Komplex von phylogenetischen Änderungen kommt als fünfte Dimension hinzu, da hier die Unterschiede und Gemeinsamkeiten von Entwicklungswegen über die Artgrenzen hinaus und über die Existenzdauer der jeweiligen Art geklärt werden müssen. Vor der Erläuterung verschiedener Hypothesen, die eine Änderung ontogenetischer Entwicklungsabläufe als Mechanismus zur evolutionären Entstehung des Vogelflügels favorisieren, muss deshalb ein Überblick zum Kenntnisstand der Embryonalentwicklung der vorderen Gliedmaßen bei Vögeln und Landwirbeltieren stehen. In den letzten Jahrzehnten wurden zahlreiche beschreibende und experimentelle Studien am Flügel des Hühnchens sowie der vorderen Extremität der Maus durchgeführt. Diese Strukturen gehören damit zu den am besten untersuchten und verstandenen Objekten der Embryologie (Welten 2005, Tickle 2006).
Morphologische Gestaltbildung
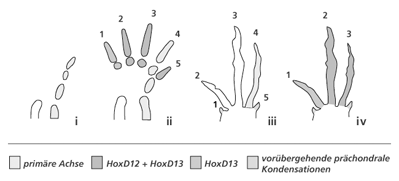 |
| Abb. 4: Die primäre Achse und prächondrale Kondensationen. a Die primäre Achse wird aus den embryonalen knorpligen Anlagen von Humerus, Ulna und dem späteren 4. Finger bei den fünffingrigen Wirbeltieren (b) gebildet. c Überträgt man diese Zählweise auf die embryonale Situation beim Vogelflügel, muss der hinterste Finger mit der Nummer 4 benannt werden. Die Existenz prächondraler Anlagen (1,5) unterstützt diese Zählweise. d Die paläontologische Zählweise belegt demgegenüber den hintersten Finger mit der Nummer 3. Das Auftreten spezieller Hox-Genexpressionsmuster während der Ontogenese soll diese Bezeichnungen rechtfertigen (vgl. Vargas & Fallon 2005). |
|
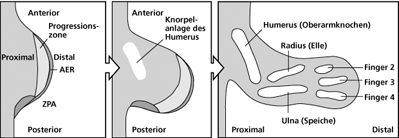
Startpunkt der Gliedmaßenentwicklung bei Wirbeltieren sind kleine Knospen („Buds“) an den seitlichen Flanken des embryonalen Körpers. Diese Knospen entstehen in einer definierten topographisch-anatomischen Region entlang der Anlagen der Wirbelsäule – den so genannten Somiten. In den Extremitätenknospen befinden sich zunächst nur unspezialisierte mesenchymale Zellen, die von einer Schicht embryonaler Haut, dem Ektoderm, bedeckt werden. Rein deskriptiv betrachtet beobachtet man im weiteren Verlauf Einwanderungen von Zellen aus dem Gebiet der Somiten und ein Auswachsen der Knospe in proximo-distaler Richtung von der Körperwand weg. Die unspezialisierten mesenchymalen und eingewanderten Zellen vermehren und differenzieren sich zu den verschiedenen die Extremität konstituierenden Zelltypen (Muskel, Bindegewebe, Knorpel etc.). Parallel dazu werden knorpelige Anlagen der Extremität in einer definierten Reihenfolge sichtbar, welche die wesentlichen Elemente des Grundbauplans vom Skelett der Wirbeltierextremität repräsentieren (Oberarm, Unterarm, Mittelhand). Nachfolgend werden die knorpeligen Anlagen der Fingerknochen sichtbar, die eine große Variabilität hinsichtlich der Anzahl der einzelnen Glieder (Phalangen) und ihrer individuellen Ausprägung bei den Wirbeltieren aufweisen (vgl. dazu Abb. 3).
Für unsere Betrachtungen von besonderer Bedeutung ist die Tatsache, dass bei fast allen fünffingrigen (pentadaktylen) Amnioten die Reihenfolge des Auftretens der ersten embryonalen Knorpelanlagen in der proximo-distalen Achse identisch ist: Humerus (Oberarmknochen) – Ulna (Unterarmknochen – Elle) – Mittelhandknochen „IV“ – Fingerknochen „IV“. Diese Reihenfolge wird als „Primäre Achse“ (Burke & Alberch 1985) bezeichnet und begründet die Festlegung der als homolog zu bezeichnenden Finger beim Artvergleich. Beim Hühnchenembryo läuft die „Primäre Achse“ durch die am weitesten posterior (hinten) gelegene Fingeranlage, die deshalb die Kennzeichnung IV erhält, und konsequenterweise werden die beiden davor gelegenen Fingeranlagen mit II und III gekennzeichnet (vgl. Abb. 4). Die Richtigkeit dieser Interpretation wird neben Mutationsexperimenten (Morgan et al. 1992) durch den Befund unterstützt, dass typische prächondrale Kondensationen (das sind Gewebestrukturen, die der Knorpelbildung vorangehen) vor (anterior) der Fingeranlage II und hinter (posterior) der Fingeranlage IV in der Flügelknospe bei Vögeln nachweisbar sind. Diese repräsentieren nach Ansicht der Entwicklungsbiologen temporäre bzw. rudimentäre Anlagen der Finger I und V (Welten 2005), die jedoch im Weiteren nicht zu vollständigen Fingern weiterentwickelt werden.
Die Diskrepanz zwischen der phylogenetischen Interpretation der paläontologischen Befundlage (siehe Abschnitt „Ableitung der Vögel von den Dinosauriern“) und die Deutung unter Nutzung der embryologischen Daten bezüglich der Identität der Fingerstrahlen bei Vögeln ist schon seit dem Beginn der evolutionären Morphologie Mitte des 19. Jahrhunderts wahrgenommen worden und wird bis heute kontrovers diskutiert. Die embryonal begründete Homologie wird von zahlreichen Forschern als sekundäre Situation bewertet, die sich erst durch phylogenetische Transformationen (siehe unten) aus der ursprünglichen embryonalen Situation bei Archosauriern entwickelt habe (z.B. Wagner & Gauthier 1999, Vargas & Fallon 2005, Wagner 2005).
Genetische und epigenetische Regulationskaskaden der raum-zeitlichen Gestaltbildung der Extremitäten
Obwohl morphologisch kaum Unterschiede innerhalb der Extremitätenknospe wahrnehmbar sind, existiert bereits auf molekularbiologischer und genetischer Ebene eine komplexe Architektur sich gegenseitig beeinflussender Kompartimente. Diese werden u.a. dadurch charakterisiert, dass zeitlich und räumlich geordnet unterschiedliche Morphogene (das sind Eiweißstoffe, deren räumliches Konzentrationsgefälle zu einer entsprechenden räumlich geordneten Spezifikation von Zellen führt) und differierende genetische Expressionsmuster innerhalb von Zellgruppen aktiv sind. Experimentell orientierte Untersuchungen führten zur Unterscheidung funktionell und regulatorisch wichtiger Areale an der Extremitätenknospe:
- „AER“ (apical ectoderm ridge), eine ektodermale Leiste, am körperfernen (distalen) Ende der Anlage von vorn (anterior) nach hinten (posterior) verlaufend,
- „PZ“ (progress zone), die Progressionzone welche mit undifferenzierten, aber genetisch vorgeprägten mesenchymalen Zellen unter der Kappe des „AER“ aufgefüllt ist, und die
- „ZPA“ (zone of polarizing activity), eine polarisierende Region am hinteren (posterioren) Rand der Extremitätenknospe, der Ort, wo später der kleine Finger bei Säugetieren entstehen wird. Eine zentrale Rolle spielt das hier produzierte Morphogen Shh (sonic hedgehog, vgl. Abb. 5).
Für alle drei raum-zeitlichen Achsen der Extremitätenentwicklung vermutet man unterschiedlich agierende Regulationssysteme. Diese rekrutieren u.a. zahlreiche Zellsignalstoffe, Wachstumsfaktoren, Morphogene und Hox-Gene der Gruppen HoxA9-13 und HoxD9-13. Aus deren geordnetem Wechselspiel entstehen die jeweils typischen Merkmalsmuster der Arme, der Hand oder des Flügels. Während für die proximo-distale und die anterio-posteriore Musterbildung relativ viel Detailwissen verfügbar ist, sind die Kenntnisse bezüglich der ventro-dorsalen Merkmalsdifferenzierung sehr gering. Allerdings – und das macht ein Verständnis der Gesamtsituation nicht einfacher – diskutieren Forscher z.T. unterschiedliche Modelle für die Strukturbildung entlang einer Achse. An dieser Stelle soll nur kurz auf die vermuteten Prozesse, die zur Differenzierung entlang der proximo-distalen Achse führen, eingegangen werden. (Im Kastentext werden zwei Modelle der anterio-posterioren Musterbildung vorgestellt).
Summerbell, Lewis und Wolpert entwarfen 1973 das „progress-zone“ Modell, das sich bis zum Ende der 1990er Jahre behaupten konnte. Danach kommt der Zeitdauer, die Zellen in der „progress zone“ (PZ) verbringen, und den während dieser Zeit auf sie einwirkenden Signalen aus der „apical ectoderm ridge“ (AER) z.B. über Morphogene der FGF-Gruppe eine positionsdeterminierende (spezifizierende) Bedeutung zu. Treten während des Wachstums die Zellen aus der „PZ“ heraus, beginnen sie sich entsprechend ihrer positionsdeterminierten Prägung zu differenzieren. Deshalb werden zuerst die Strukturen des Oberarmknorpels, dann des Unterarmknorpels und am Ende die der Fingerknorpel sichtbar. Aktuelle experimentelle Ergebnisse konnten demgegenüber zeigen, dass entgegen dem „progress zone“-Modell bereits auf der frühesten Stufe der Existenz der Extremitätenknospe das proximo-distale Muster in verschiedenen Zelldomänen durch einen unbekannten Mechanismus festgelegt sein muss. Der Einfluss des vom „AER“ ausgehenden Morphogens FGF dient danach dazu, diese spezifizierten Muster über einen größeren Raum auszudehnen. Und diese Ausweitung oder Übertragung der vorliegenden Positionswerte geschieht zeitversetzt. D.h. die proximalen, körpernahen Anteile werden früher expandiert und damit eher sichtbar als die der distalen Extremitätenstrukturen.
Dieses „early specification and progressiv determination model“ (Dudley et al. 2002) wurde von Richardson et al. (2004) weiter verfeinert. In der Zelldomäne, aus der die distalen Stukturen der Extremität – also die Mittelhandknochen und Finger – hervorgehen, wird erst spät ein zyklisch agierendes Programm wirksam, welches die Segmentation (Unterteilung) der Finger in die charakteristische Anzahl ihrer Glieder steuert. Die Anzahl der Finger und die Zahl der einzelnen Fingerglieder wäre danach noch nicht in der Extremitätenknospe spezifiziert. Die späte Determinierung der Fingerstrukturen wird durch den Befund gestützt, dass die typische Morphologie der einzelnen Finger von mesenchymalen Zonen bestimmt wird, welche zwischen den Fingern gelegen sind (IDM – interdigital mesenchyme, Dahn & Fallon 2000). Manipulationen am IDM beim Hühnchen führten zur isolierten Änderung der jeweiligen Fingermorphologie in die eines anderen Strahls (z.B. Finger II entwickelte die Merkmale von Finger III).
Das heute dominierende „early specification model“ der proximo-distalen Entwicklung der Extremität weist Parallelen zum Morphogen-Gradienten Modell auf (siehe Kastentext 1), welches für die anterio-posteriore Musterbildung der Handentwicklung diskutiert wird.
Unbestritten ist die Tatsache, dass zwei Hauptmechanismen – Gradienten von Morphogenen und die Zeitdauer ihrer lokalen Wirkung – bei der Differenzierung in beiden Achsen wirksam sind. Wie angedeutet bleibt noch unklar, ob diesen Mechanismen eine ebenso dominierende Rolle bei der Ausbildung der dorso-ventralen Differenzierung zukommt (Tickle 2006). Nachgewiesen wurde bislang ein fördernder Einfluss des Wachstumsfaktors Wnt 7a auf die Prägung von Strukturen des Handrückens und des Transkriptionsfaktors En1 auf die der Handfläche in Experimenten an der Maus (Cygan 1997).
|
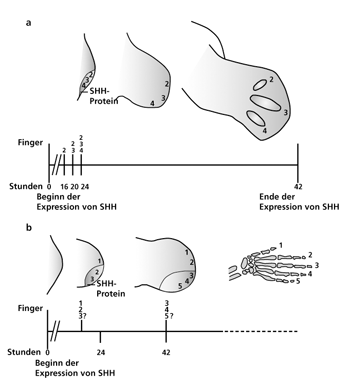
A. Morphogen-Gradienten–Modell beim Hühnchen. Die Konzentration des Morphogens „sonic hedgehog“ (SHH) spezifiziert Zellgruppen zeitlich gestaffelt mit Positionswerten für jeden einzelnen Fingerstrahl des Flügels in einem frühen embryonalen Stadium (16-24 Stunden nach Beginn der Expression von SHH). Aus den so vorgeprägten Zellen entwickeln sich in einer späteren Phase die jeweiligen Fingerstrahlen. Der Zeitstrahl dokumentiert die Dauer der SHH-Expression. 24 Stunden nachdem die Positionswerte fixiert sind, beginnen sich die Anlagen der Finger zu entwickeln.
B. Weiterentwickeltes Morphogen-Gradienten–Modell am Beispiel der Maus. Die Konzentration und die Länge der Wirkung von SHH auf einzelne Zellgruppen bestimmt deren Prägung mit spezifischen Positionswerten, die dann die Entwicklung der jeweiligen Finger festlegen.
Der erste Finger (Daumen) entwickelt sich nach diesem Modell unabhängig vom Einfluss des SHH, der Positionswert für Finger wird von der Konzentration von SHH in einer frühen Phase bestimmt. Die Positionswerte von Finger drei werden bestimmt von der Konzentration und der Dauer der SHH-Wirkung, die der Finger 4 und 5 allein von der Dauer der SHH-Wirkung. Zusätzlich kommt der Entwicklungsphase eine wichtige Rolle zu; die hinteren (posterioren) Finger werden in späteren Phasen als die vorderen spezifiziert.
|
Bedeutung der Hox-Gene
Abschließend noch einige Ausführungen zur Bedeutung der Hox-Gene im Rahmen der Extremitätenentwicklung, da diese in der phylogenetischen Argumentation (siehe unten) gegenwärtig eine Schlüsselrolle einnehmen.
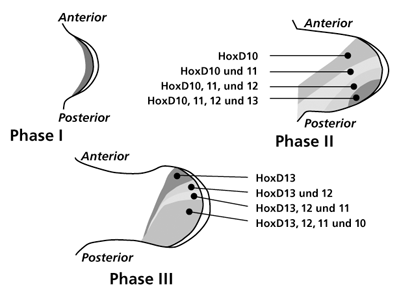 |
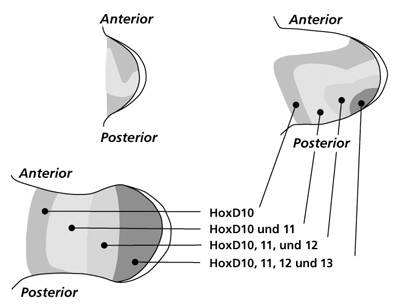
| Abb. 6: Schematische Darstellung der HoxD-Gen-Cluster Expressionsmuster in drei aufeinander folgenden Entwicklungsstadien der Extremitätenknospe bei Wirbeltieren. Man beachte den fast spiegelbildlichen Wandel der Expressionsmuster in Phase II und III. Eine Homologisierung von Zellgruppen mit einem speziellen HoxD-Gen-Expressionsmuster mit morphologischen Strukturen ist nicht möglich. Nach www.mun.ca/...; www.nature.com/... |
|
Hox-Gene codieren für Transkriptionsfaktoren, die ihrerseits nachgeschaltete Regulationsgene und Strukturgene beeinflussen, aber auch selbst für Strukturproteine codieren können (Pearson et al. 2005). Diese besonderen Gene sind auf vier Hox-Gen-Cluster bei Vögeln und Säugetieren verteilt (A, B, C, D) und besitzen clusterabhängig eine unterschiedliche Bedeutung für die Extremitätenentwicklung. Die Gene 9-13 der Cluster A und D (HoxA 9-13 und HoxD 9-13) sind unverzichtbar für eine normale Ausprägung der Gliedmaßen, die homologen Gene der Cluster B und C dagegen nicht (Dlugaszewska et al. 2006). Die Vermutung, dass einzelne Hox-Gene für spezifische Strukturen allein verantwortlich sind (also HoxA 10 für Oberarmknochen, HoxA 10 und 11 für Unterarmknochen usw.) hat sich nicht bestätigt. Es existiert kein starres homologes Abbild zwischen den Expressionsmustern der beteiligten Hox-Gene und den morphologischen Entsprechungen der Extremitätenentwicklung. Ein Beispiel: In den einzelnen Phasen der Entwicklung breiten sich wellenartig mehrere, zum Teil reverse (spiegelbildliche) Expressionsmuster des HoxD-Clusters ausgehend von der „ZPA“ über die Extremitätenknospe aus, ohne eine enge Korrelation zu einer bestimmten Achse aufzuweisen (Abb. 6). Das Expressionsmuster der HoxA-Gene zeigt dagegen eine auffällige Anordnung und Ausbreitung entlang der proximo-distalen Achse (HoxA 10, HoxA 11, HoxA 12, HoxA 13), die mit der späteren anatomischen Unterteilung eher zu korrelieren scheint (Abb. 7).
Im Hinblick auf die lokal typische Spezifikation der Finger erscheint die Wirkung der Hox-Gene von untergeordneter Bedeutung. Tickle (2006) vermutet, dass in der letzten Phase durch die HoxD-Gen-Expression Positionswerte vermittelt werden könnten, die den Fingeranlagen ihre spezifische Identität vermitteln. Experimentelle Untersuchungen lassen jedoch Zweifel an dieser Interpretation aufkommen. Manipulationen an den Fingeranlagen, in deren Ergebnis die Identität von Finger II zu Finger III oder von Finger I zu Finger II verändert wurden, zeigten keine Änderungen des vorangehenden bekannten normalen Expressionsmusters von HoxD 10-12 (Tickle 2006).
Welche konkrete Rolle den Hox-Genen und anderen „Masterkontrollgenen“ im Rahmen der vierdimensionalen Entwicklungsvorgänge der Extremität bei Wirbeltieren zukommt und wie entscheidend ihr Beitrag bei der Festlegung der Strukturbildungen in den jeweiligen Achsen ist, konnte aufgrund ihrer komplexen und überlappenden Wirkungen bislang noch nicht schlüssig aufgeklärt werden. Ohne dieses Wissen fehlt aber die fundamentale Voraussetzung dafür, den vermuteten phylogenetischen Wandel auf kausaler Ebene auch nur annähernd plausibel begründen zu können. Für die Homologiediskussion ergeben sich aus den Erkenntnissen der Ontogenese zusätzliche methodische Schwierigkeiten. Nach Galis (2005) und Feduccia (2005) sprechen die netzartig agierenden Teilprozesse der Genexpression von Hox-Genen sowie ihr zeitlich und räumlich variierendes Expressionsmuster dagegen, einzelne genetische Domänen einer Entwicklungsphase als Marker für morphologische Strukturen (z.B. HoxD-13 zu Finger I bei Vargas 2005) und damit als Basis für Homologiediskussionen zu nutzen.
|
|
4. Wie wird aus der Dinosaurierhand ein Vogelflügel?
Ontogenetischer – anatomisch/paläontologischer Datenvergleich und resultierende Möglichkeiten ihrer homologen Korrelation
Überblick: Die Evolutionsbiologie ist herausgefordert, neben Antworten auf die stammesgeschichtliche Herleitung der Vögel auf der Grundlage von Homologien die kausalen Hintergründe phylogenetischen Wandels plausibel zu machen. Die vorhandenen Kenntnisse aus der Embryologie, Anatomie und Paläontologie erlauben unterschiedliche Homologisierungen und damit die Vorgabe mehrerer stammesgeschichtlicher Szenarien der Flügelentwicklung. Es fehlt ein methodisch sicheres Kriterium, um aus den Homologiezuweisungen auf der Basis embryologischer und paläontologischer Daten die phylogenetisch relevanten zu extrahieren. Keiner der vorgestellten Ansätze kann deshalb für sich ein höheres Maß an Plausibilität beanspruchen als ein anderer. Dieses Dilemma könnte behoben werden, wenn Mechanismen entdeckt würden, die den vermuteten phylogenetischen Wandel auf genetischer und molekularbiologischer Ebene wahrscheinlich machen. Je nach favorisiertem Homologieansatz diskutiert man mehrere spekulative kausale Szenarien. Vier hypothetische Lösungsansätze stehen dabei im Vordergrund: „primary axis shift“-Hypothese, „frame shift“-Hypothese, bilaterale Reduktion, primäre Polydaktylie. Die „frame shift“-Hypothese erfährt gegenwärtig den meisten Zuspruch. Sämtliche kausalen Erklärungsversuche zur Entstehung des Grundbauplans des Vogelflügels auf der Grundlage favorisierter Mechanismen der Modernen Synthetischen Evolutionstheorie (Kutschera 2006) bzw. der Evolutionären Entwicklungsbiologie (Evo-Devo) sind hypothetisch und als spekulative Extrapolationen von Details des verfügbaren, äußerst lückenhaften embryologischen bzw. molekulargenetischen Wissens zu werten.
Die Homologie zwischen dem Vogelflügel und der vorderen Extremität von Dinosauriern und ihre kausale Begründung bleibt unter phylogenetischer Perspektive auch nach 150 Jahren Evolutionsforschung unklar.
Wagner (2005) resümiert, dass die Klärung der Homologie der Fingerstrahlen des Flügels bei Vögeln ein ungelöstes Problem seit dem Beginn der evolutionären Morphologie Mitte des 19. Jahrhunderts sei. Als Kernproblem dafür macht er die hartnäckig existierende Diskrepanz zwischen den anatomisch/paläontologischen und embryologischen Befunden aus, die für sich genommen jeweils eine andere phylogenetische Homologie erfordern. Im Folgenden werden die wichtigsten hypothetischen Ansätze zur Korrelation der vorliegenden Datensätze vorgestellt, die in den letzten zwei Jahrzehnten entworfen und kontrovers diskutiert wurden (vgl. dazu Abb. 8).
Zum besseren Verständnis der jeweiligen Entwürfe erfolgt deren Darstellung nach einem vergleichbaren Schema hinsichtlich der Identifikation der embryonalen Anlagen, hier mit dem Buchstaben C gekennzeichnet (CI-CV, CI = embryonale Anlage des Daumens, CV embryonale Anlage des 5. Fingers) und der ausgebildeten Finger, die den Buchstaben D erhalten (Finger DI-DV, DI=Daumen, DV=5. Finger). In einer Variante wird noch ein Präpollex (ein vor dem Daumen gelegener Finger) eingebunden, der entweder mit CP oder DP gekennzeichnet wird.
Zuerst werden für jeden Erklärungsansatz die angenommenen phylogenetischen Vorfahren benannt, danach die vermuteten Prozesse des phylogenetischen Wandels mit den möglichen hypothetischen Evolutionsmechanismen. Das abgeleitete Ergebnis wird mit dem anatomisch-embryonalen Befund der Vogelflügelentwicklung korreliert und eine mögliche Homologie begründet. Abschließend sind jeweils einige Argumente, welche für die aufgestellte Homologie sprechen und die wesentlichen Kritikpunkte, die sie in Frage stellen, angeführt (vgl. dazu Abb. 8).
1. „axis shift“-Hypothese – ASH (Abb. 8.1): Verlagerung der primären embryonalen Achse vom IV. auf den III. Strahl (Shubin 1994; Chatterjee 1998; Garner & Thomas 1998)
Basale Archosaurier mit der Fingerformel (DI, DII, DIII, DIV, DV) entwickeln sich zu Theropoden mit drei Fingern (DI, DII, DIII) durch Reduktion von CIV, CV und DIV, DV. Damit parallel bzw. in der weiteren Entwicklung in Richtung zu den Vögeln erfolgte ein Versatz der primären embryonalen Achse („axis shift“) von der Fingeranlage CIV auf CIII durch ein anderes „Timing“ im Erscheinen der prächondralen Anlagen (CIII erscheint jetzt als erste Anlage). Die drei Fingerstrahlen der Theropoden und des Vogelflügels sind deshalb homolog zu DI, DII, DIII der übrigen Landwirbeltiere (Abb. 8.1).
Das genetische Expressionsmuster von HoxD12 und HoxD13 in den Fingeranlagen der ersten zwei Flügelstrahlen und die paläontologisch begründete Theropodenabstammung der Vögel sind damit kompatibel.
 |
| Abb. 8: Schematische Darstellung der vier Hypothesen zur Ableitung der Vogelhand. |
|
Kritik (vgl. Wagner & Gauthier 1999, Wagner 2005, Feduccia 2005, Vargas & Fallon 2005 a, b): Selektionsdrücke, die eine solche phylogenetische Änderung „erzwingen“, sind bislang nicht einsichtig. Ebenso ist der molekulargenetische Mechanismus eines solchen „Shifts“ unbekannt. Die postulierte Änderung der primären Achsen dürfte sich zudem nur auf die Flügel beschränken und die untere Extremität nicht beeinflussen. Experimentelle Untersuchungen dagegen zeigen, dass bei grundlegenden genetischen Veränderungen Hand und Fuß immer gleichermaßen betroffen sind. Bei Vogelembryonen werden regelhaft drei anteriore (vordere) vor und eine posteriore (hintere) prächondrale Fingeranlage hinter der primären embryonalen Achse gefunden (s. Abb. 8)!
Dieser Befund deckt sich mit der Situation bei fast allen anderen Amnioten und stützt die Interpretation, dass die primäre embryonale Achse auch bei Vögeln durch die vierte Fingeranlage verläuft.
2. „frame shift“-Hypothese – FSH (Abb. 8.2): Komplexe Verschiebung der morphologischen Identitäten von drei Fingern nach vorn (Wagner & Gauthier 1999, Wagner 2005, Vargas & Fallon 2005 a, b)
Basale Archosaurier mit der Fingerformel (DI, DII, DIII, DIV, DV) entwickeln sich zu Theropoden mit drei Fingern (DI, DII, DIII) durch Reduktion von DIV und DV, wobei als embryonale Anlage CIV erhalten bleiben muss. Damit parallel bzw. in der weiteren Entwicklung in Richtung zu den Vögeln erfolgte (im Unterschied zur ASH) eine homeotische Transformation („frame shift“) der Determinierungsmuster in den prächondralen Fingeranlagen dergestalt, dass aus CI kein adulter Finger mehr entsteht und aus CII, CIII und CIV sich Finger mit der anatomischen Identität von DI, DII und DIII entwickelten (und nicht wie sonst im Normalfall DII, DIII, DIV). Die embryonalen Fingeranlagen sind homolog zu CII, CIII und CIV der übrigen Wirbeltiere, die adulten Fingerstrahlen des Vogelflügels dagegen homolog zu DI, DII, DIII der Theropoden (Abb. 8.2). Die primäre embryonale Achse läuft durch CIV wie bei allen anderen Amnioten, aus der embryonalen Anlage des 4. Fingers (CIV) bildet sich dann durch „frame shift“ ein Finger mit den Merkmalen von DIII.
Das genetische Expressionsmuster von HoxD12 und HoxD13 in den Fingeranlagen der ersten zwei Flügelstrahlen und die paläontologisch begründete Theropodenabstammung der Vögel sind damit kompatibel.
Die vermutete homeotische Transformation der Finger als kausaler Hintergrund ähnelt jenem, der auch bei der Entwicklung der Wirbelkörper angenommen wird (vgl. Ullrich 2005).
Kritik (Galis et al. 2003, Feduccia 2005, Welten et al.): Selektionsdrücke, die eine solche phylogenetische Änderung „erzwingen“ sind auch für dieses Szenario nicht einsichtig. Die postulierte homeotische Transformation beschränkt sich nur auf die Flügelstrahlen. Auch hier gilt nach experimentellen Erfahrungen, dass bei grundlegenden genetischen Veränderungen Hand und Fuß gleichermaßen betroffen sind. Die FSH benötigt viele Hilfsannahmen: Sie setzt voraus, dass bei den Theropoden trotz Reduktion der Finger DIV und DV embryonal CI bis CIV angelegt blieben und durch CIV die embryonale Achse weiterhin verlief. Weiterhin behielt danach die vorhandene CIV Anlage bei den Vorläufern zu den Vögeln die Fähigeit, Finger zu bilden; diese Fähigkeit wurde jedoch erst beim Schritt zu den Vögeln reaktiviert und zwar so, dass nun aus CIV die DIII-Identität wie bei Archaeopteryx entsteht.
Diese Hypothese setzt weiterhin voraus, dass auch die korrespondierenden Änderungen im Bereich der Handwurzel parallel erfolgten. Es gibt aber noch keine Untersuchungen über den Einfluss von Hox-Genen und von homeotischen Mutationen auf die Handwurzel. Der molekulargenetische Mechanismus der vermuteten homeotischen Transformation zur Veränderung des vorliegenden Phänotyps ist unbekannt und der postulierte homeotische Shift erscheint einzigartig innerhalb des breiten Spektrums der embryonalen Entwicklungswege der Amnioten. Es existieren keine paläontologischen Befunde bei den dreifingrigen Theropoden, die auf eine rudimentäre Anlage von CIV hinweisen.
3. Bilaterale Reduktion des ersten und fünften Fingers und sekundäre Umgestaltung der Fingerstrahlen (Abb. 8.3, Kundrát et al. 2002, Galis et al. 2003, Feduccia 2005)
Basale Archosaurier mit der Fingerformel (DI, DII, DIII, DIV, DV) entwickeln sich zu Formen mit drei Fingern (DII, DIII, DIV) durch pyramidale Reduktion von DI und DV (Abb. 8.3). Entweder über die Theropoden oder auf direktem Wege erfahren die verbleibenden Fingerstrahlen der hypothetischen Vorfahren in der weiteren Entwicklung in Richtung zu den Vögeln einen morphologischen Umbau hinsichtlich der Gestalt, der Anzahl und Größe der einzelnen Fingerglieder (z.T. durch Verlust bzw. Zugewinn von Fingergliedern), in dessen Ergebnis sich die Phänotypen der Finger von Theropoden und von Archaeopteryx annäherten. Sind die Vögel die direkten Nachkommen der Theropoden, hat sich dieser Prozess bereits in seinen Grundzügen auf dem Weg von Archosauriern zu den Theropoden vollzogen. Genauso wahrscheinlich ist es, dass Theropoden und Vögel Schwesterngruppen sind und damit die morphologische Identität ihrer Finger eine Konvergenzentwicklung darstellt.
Die adulten Fingerstrahlen des Vogelflügels sind homolog zu DII, DIII, DIV der basalen Archosaurier und homolog zu den Fingern der Theropoden und würden damit die Finger DII-DIV repräsentieren.
Das Erscheinungsbild der Finger bei Wirbeltieren ist hinsichtlich der morphologischen Differenzierung und der Anzahl der einzelnen Fingerglieder äußerst variabel und wird ontogenetisch relativ spät determiniert (s.o.). Vergleichende phylogenetische Studien führen zu der Vermutung, dass es ein allgemeines evolutionäres Muster ist, dass der erste Finger verloren geht oder reduziert wird und dann der 5. Finger (Shapiro et al. 2003).
Ontogenetisch gelang der Nachweis von 5 Fingeranlagen am Flügel (prächondrale Kondensationen) mit einem typischen Expressionsmuster des Sox9-Gens (rudimentäre Anlagen des I. und V. Fingers), obwohl sich später nur drei Strahlen entwickeln.
Galis et al. (2003) konnten zeigen, dass in sehr frühen Phasen in der Entwicklung des ersten Strahls auch die Expression von HoxD12 nachweisbar ist.
Kritik (Wagner & Gauthier 1999, Wagner 2005, Vargas & Fallon 2005 a,b, Welten et al. 2005): Das späte genetische Expressionsmuster von HoxD12 und HoxD13 in den Fingeranlagen DII und D III widerspricht dieser Hypothese (s.o.). Es sind keine Selektionsdrücke und keine Mechanismen bekannt, die eine isolierte Reduktion von DI und DV verständlich machen würden. Die paläontologisch begründete Einordnung der Theropoden müsste revidiert werden, da deren Fingerformel nicht mehr auf der Reduktion des IV. und V. Fingers der Hand begründet werden kann. Nach dieser Hypothese beruht die morphologische Identität des ersten Strahls von Archaeopteryx und des ersten Strahls bei Maniraptoren auf Konvergenz, auch hinsichtlich der Anzahl der einzelnen Fingerglieder und der Klaue. Es fehlen für dieses Szenario paläontologische Belege (z.B. intermediäre Baupläne oder ein früher Theropode mit reduziertem, vor dem eigentlich ersten Strahl gelegenem Fingerrudiment).
4. Abstammung der Vögel und der Theropoden von sechs- oder vielfingrigen (hexadaktylen/polydaktylen) Archosauriern (Abb. 8.4) (Welten et al. 2005)
Alle Vögel und Theropoden leiten sich von sechs- und mehrfingrigen Archosauriern ab, welche vor dem 1. Finger mindestens noch einen zusätzlichen Strahl (Präpollex) im embryonalen und adulten Stadium besaßen (CP, DP). Beim Übergang zu den Theropoden und Vögeln wurden der vorderste Strahl (P) sowie DIV und DV rückgebildet (analog wie bei der Hypothese der pyramidalen Reduktion).
Beim Übergang zu den fünffingrigen (pentadaktylen) Archosauriern (von welchen sich die heutigen Reptilien und Säugetiere ableiten) wurde aber der Präpollex (P) beibehalten und DV reduziert.
Die weitere morphologische Ausprägung und Remodellierung der Fingerstrahlen bei Theropoden und Vögeln erfolgt auf dem selben Weg, wie er bei der Hypothese der pyramidalen Reduktion bereits dargestellt wurde.
Die Homologie von DI-DIII bei Theropoden und Vögeln lässt sich im Hinblick auf die ontogenetische (Muster der prächondralen Anlagen) und paläontologische Datenlage aufrechterhalten. Es gelingt weiterhin auch eine befriedigende Deutung der Situation im Hinblick auf die heutigen Reptilien und Säugetiere (eigentliche Fingerformel: P – I – II – III – IV, primäre Achse verläuft durch die „III“-Anlage).
Kritik (Welten et. al. 2005): Selektionsdrücke, die eine solche phylogenetische Änderung „erzwingen“, sind wiederum nicht einsichtig. Ebenso ist der molekulargenetische Mechanismus einer pyramidalen Reduktion unbekannt. Das identische späte genetische Expressionsmuster von HoxD12 und HoxD13 in den ersten Fingeranlagen beim Hühnchen und der Maus widerspricht dieser Hypothese (s.o.).
Es gibt keine ontogenetischen Daten von Theropoden, um die Existenz eines Präpollex (CP) zu stützen. Ebenso kennt man bislang keinen paläontologischen Fund eines adulten Archosauriers mit sechs Fingern (wohl aber Formen mit mehr als sieben) oder mit einer rudimentären vorderen Fingeranlage bei frühen Theropoden.
Bei der ontogenetischen Entwicklung des Flügels und des Fußes bei Vögeln konnten bislang immer nur fünf und keine sechs digitalen Domänen des für Fingeranlagen spezifischen Sox9-Gens nachgewiesen werden.
Zusammenfassung
Alle vier hypothetischen Ansätze verfolgen das Ziel, die Diskrepanz zwischen den verfügbaren anatomisch/paläontologischen und ontogenetischen Befunden zu überwinden. Die „frame shift“- und die „axis shift“-Hypothesen basieren auf der Vorgabe, dass durch Änderungen embryonaler Mechanismen (homeotische Transformation, Änderung des „Timings“ von Entwicklungsschritten) und darauf aufbauender adaptiver Änderungen der Fingermorphologie der Wandel von der dreifingrigen Hand der Theropoden zum dreistrahligen Flügel der Vögel möglich wurde. Für die hier genutzten Evolutionsmechanismen, die auf einer Änderung ontogenetischer Prozesse basieren („EvoDevo“), gibt es jedoch keine experimentelle Basis bzw. ein theoretisches Modell, das mit dem bekannten ontogenetischen Wissen verträglich wäre.
Die Hypothesen der pyramidalen Reduktion und ihre Abwandlung unter der Vorgabe einer sechs- oder mehrfingrigen Ausgangsform favorisieren kausal nicht näher begründete Reduktionstendenzen der vorderen und hinteren Finger und ausgeprägte Remodellierungen der verbliebenen Finger. Die verbleibenden Konflikte mit paläontologischen und ontogenetischen Befunden sowie die verstärkte Inanspruchnahme von Konvergenzentwicklungen sind Kernpunkte der diesbezüglich vorgetragenen Kritik.
Allen Harmonisierungsversuchen ist darüber hinaus gemeinsam, dass von den Autoren zu den Selektionsdrücken, welche die dramatischen Umbauten kanalisiert haben, keinerlei plausible Angaben gemacht werden können.
Mittels wissenschaftlicher Kriterien kann keine der aufgeführten Hypothesen bislang eine Überlegenheit gegenüber den anderen nachweisen oder für sich eine besondere Plausibilität beanspruchen. Neben den oben diskutierten Varianten wäre es genauso berechtigt zu behaupten, Vögel und Theropoden gehörten nicht zu einem gemeinsamen „Clade“ (Abstammungszweig) oder die Nutzung der Fingermorphologie zur Klärung der Homologie sei nicht sinnvoll oder die ontogenetisch nachweisbare Struktur (CI) bei Vögeln habe nichts mit einer Fingeranlage zu tun (Welten 2005).
 |
Zu den Selektionsdrücken,
die die dramatischen
Umbauten kanalisiert haben,
gibt es keine plausiblen Argumente. |
 |
|
Die in der wissenschaftlichen und populärwissenschaftlichen Literatur vorherrschende Favorisierung der „frame shift“ Hypothese, welche die Theropodenabstammung der Vögel unterstützt, findet bei Wagner (2005) eine eigenartige sowie aufschlussreiche Begründung. Auf die Kritik der von ihm vertretenen „frame-shift“ Hypothese durch Feduccia (1999) und Galis et. al. (2003) schreibt er:
„Beide Argumente stellen wahre Fakten dar, nämlich dass unser Wissen über die Mechanismen inkomplett ist und dass es keine kausale Erklärung dafür gibt, was wir in der Evolution der Vögel sehen. Und es ist wahr, dass kein Szenario für die Anpassungen existiert, welches die Differenzen in der Fingerentwicklung zwischen Vögeln und Alligatoren erklärt. Aber diese Fakten sind eindeutig nebensächlich, die Frage, wie die Finger und die Ontogenese der Finger bei den Theropoden evolvierten, ist eine Frage, die unabhängig davon betrachtet werden muss, ob wir das Wissen oder die Werkzeuge besitzen um herauszufinden, welche Mechanismen den Wandel verursacht haben. Ein Scheitern, das Geschehene zu erklären, hat keine Auswirkung darauf, ob ein Ereignis sich vollzogen hat oder nicht. Dieses Scheitern reflektiert nur die Grenzen unseres gegenwärtigen Wissens. Ein historischer Fakt kann wahr sein, ob wir ihn erklären können oder nicht.“ (Hervorhebungen nicht im Original)
Unter Berücksichtigung der Situation, dass die oben diskutierten Diskrepanzen sich bereits aus der Betrachtung nur eines Teilbereiches der vielfältigen und ambivalenten Diskussionen zur Herleitung der Vögel ergeben (vgl. dazu Junker 2005), erhält diese Aussage Wagners besonderes Gewicht. Die Evolution der Vögel ist danach kein wissenschaftlich belegtes Faktum, sondern eine postulierte Vorgabe für den Interpretationsrahmen, in dem alle Daten der molekulargenetischen, anatomischen, paläontologischen und ontogenetischen Forschung zu integrieren sind – ungeachtet ihres tatsächlichen Erklärungspotentials.
|
|
5. Das fundamentale Problem der Evolutionsbiologie: „Wann ist Homologie tatsächlich Homologie?“
In einem bemerkenswerten Artikel stellen die Autoren Wray und Abouheif (1998) diese Frage unter dem Eindruck der zunehmenden Konflikte zwischen molekulargenetischen und anatomisch/ paläontologischen Daten in der Homologiedebatte. Die Berücksichtigung ontogenetischer Daten verschärft nach Richardson et al. (2003) die aufgeworfene Problematik, weshalb neue methodische Ansätze in der vergleichenden Forschung und eine andere theoretische Basis für die Interpretation der nachgewiesenen Ähnlichkeiten erforderlich sind (vgl. Ullrich 2005).
Die allgemein am weitesten akzeptierte Definition für „Homologie“ integriert neben der deskriptiv erfassten morphologischen Identität eines Merkmals die Vorgabe, dass das betreffende Merkmal zweier Organismen auch bei einem gemeinsamen Vorfahren präsent ist. „Da die Vorfahren aber äußerst selten für eine direkte Untersuchung verfügbar sind, stellt Homologie praktisch eher eine Hypothese über die evolutionäre Geschichte als ein direktes Forschungsergebnis dar“ (Wray & Abouheif 1998).
|
„Homologie stellt eher eine Hypothese
über die evolutionäre Geschichte als ein
direktes Forschungsergebnis dar.“ |
|
|
Das Beispiel der in diesem Artikel dargestellten Debatte zur Homologie des Vogelflügels bestätigt die von diesen Autoren angeführte methodische Unzulänglichkeit des gängigen Homologieverständnisses zur Klärung phylogenetischer Fragestellungen. (Weitere im gleichen Sinn bereits diskutierte Beispiele siehe bei Ullrich 1994, 2005, 2006).
G. de Beer (1971) zählt zu den ersten, die darauf hingewiesen haben, dass aufgrund ihrer Sequenzen identische, „homologe“ Gene nicht notwendigerweise für homologe Strukturen codieren und dass homologe Strukturen nicht notwendigerweise von homologen Genen codiert werden müssen.
„Das Expressionsmuster von Genen ist mit Sicherheit keine über alle Zweifel erhabene Vorgabe zur Bestimmung von Homologien morphologischer Strukturen. Das einfache Faktum, dass Regulationsgene zahlreiche Expressionsdomänen besitzen und vielfältige Funktionen in den einzelnen Organismen wahrnehmen, machen eine solche oberflächliche Schlussfolgerung absurd“ (Wray & Abouheif 1998).
In diesen Fällen stellt eine begründbare Homologie für eine bestimmte biologische Organisationsebene (Erbgut, ontogenetischer Entwicklungsweg, ausgebildete Struktur) kein durchgängiges und auf andere Ebenen übertragbares gültiges Phänomen dar. Die bereits im evolutionären Homologiebegriff vorgegebene hypothetische Komponente einer gemeinsamen Abstammung wird durch diese Dissoziation verstärkt und schwächt damit dessen methodische Relevanz.
Die wissenschaftliche Klärung der homologen Beziehungen zwischen den Fingerstrahlen an Vogelflügeln und denen bei anderen Landlebewesen (insbesondere Echsen und Säugetiere) und ihre kausale Begründung bleibt eine der größten Herausforderungen für die Evolutionsforschung.
Das ontogenetische und phylogenetische Modellobjekt „Flügelentwicklung“ steht exemplarisch für die grundsätzlichen Probleme und Grenzen der evolutionsgeleiteten Hypothesenbildung sowie darauf aufbauender Homologiebetrachtungen. Die in der traditionellen Evolutionsbiologie vorhandenen Erklärungsmängel lassen sich nach Stotz (2005) in zwei Kategorien unterteilen: „a, Eine Theorie erklärt ein von ihr erkanntes Problem entweder nur unvollständig oder sogar unangemessen; b, eine Theorie hat ein fundamentales Problem nicht als solches erkannt. Die Übergänge zwischen diesen beiden Arten sind allerdings fließend“ (S. 349).
Es bleibt abzuwarten, ob zukünftig die evolutionäre Entwicklungsbiologie (Evo-Devo) auf die zahlreichen offenen Fragen der Evolutionsbiologie Antworten geben kann. Vielleicht dienen ihre Ergebnisse auch dazu, bisher verdrängte und fundamentale Probleme der Evolutionsbiologie zu vertiefen und neue Leitideen in der Biologie einzufordern.
|
Literatur
- Bardeleben K (1889)
- On the präpollex und prähallux, with observations on the carpus of Theriodesmus hylarchus. Proc. Zool. Soc. Lond. 1889, 259-262.
- Burke AC & Alberch P (1985)
- The development and homology of the chelonian carpus. J. Morphol. 186, 119-131.
- Chatterjee S (1998)
- Counting the fingers of birds and dinosaurs. Science 280, 355a.
- Cygan JA et al. (1997)
- Novel regulatory interactions revealed by studies of murine limb pattern in Wnt-7a and En-1 mutants. Development 124, 5021-5032
- Dahn RD & Fallon JF (2000)
- Interdigital regulation of digit identity and homeotic transformation by modulated BMB signaling. Science 289, 438-441.
- de Beer GR (1971)
- Homology: an Unsolved Problem. Oxford.
- Dlugaszewska B et al. (2006)
- Breakpoints around the HoxD cluster various limb malformations. J. Med. Genet. 43, 111-118.
- Dudley AT, Ros MA &Tabin CJ (2002)
- A re-examination of proximodistal patterning during vertebrate limb development. Nature 418, 539-544.
- Feduccia A, Lingham-Soliar T & Hinchiffle JR (2005)
- Do Feathered Dinosaurs Exist? Testing the hypothesis on Neontological and Paleontological Evidence. J. Morph. 266, 1-42.
- Galis F, Kundrát M & Sinervo B (2003)
- An old controversy solved. Bird embryos have five fingers. Trends Ecol. Evol. 18, 7-9.
- Galis F, Kundrát M et al. (2005)
- Hox genes, digit identities and the theropod/bird transition. J. Exp. Zool. (Mol. Dev. Evol.) 304B, 198-205.
- Garner JP & Thomas ALR (2004)
- Counting the fingers of birds and dinosaurs. Science 280, 355.
- Hecht MK (1985)
- The biological significance of Archaeopteryx. In: Hecht MK et al. (eds) The beginning of birds. Proceedings of the International Archaeopteryx Conference Eichstätt, pp 149-160.
- Holmgreen N (1955)
- Studies on the phylogeny of birds. Acta Zool. 36, 243–328.
- Junker R (2005)
- Der Ursprung der Vögel – ein update. Stud. Int. J. 12, 51-57.
- Kundrát M, Seichert V, Russel AP & Smetana K (2002)
- Pentadactyl pattern of the avian wing autopodium an pyramid reduction hypothesis. J. Exp. Zool. (Mol. Dev. Evol.) 294, 152-159.
- Kutschera U (2006)
- Evolutionsbiologie. 2. Aufl. Stuttgart.
- Moinar RE (1985)
- Alternatives to Archaeopteryx: a survey of proposed early or ancestral birds. In: Hecht MK et al. (eds) The beginning of birds. Proceedings of the International Archaeopteryx Conference Eichstätt, pp 209-217.
- Morgan BA et al. (1992)
- Targeted misexpression of Hox- 4.6 in the avian limb bud causes apparent homeotic transformations. Nature 358, 236-239.
- Paul GS (2002)
- Dinosaurs of the Air. The evolution and Loss of Flight in Dinosaurs and Birds. Biol. Rev. 73, 1-42.
- Pearson CP, Lemons D & McGinnis W (2005)
- Modulate Hox Gene Functions during animal Body Patterning. Nature Rev. Gen. 6, 893-904.
- Peters DS (2002)
- Anagenesis of Early Birds reconsidered. Senckenbergiana lethaea 82, 347-354.
- Richardson MK & Verbeek FJ (2003)
- New directions in comparative embryology and the nature of developmental characters. Animal Biology 53, 303-311.
- Richardson MK, Jeffery JE & Tabin CJ (2004)
- Proximodistal patterning of the limb: insights from evolutionary Morphology. Evol. Dev. 6, 1-5.
- Shapiro MD, Hanken J & Rosenthal N (2003)
- Developmental basis of evolutionary digit loss in the australian lizard Hemiergis. J. Exp. Zool. (Mol. Dev. Evol.) 297B, 48-56.
- Shubin NH (1994)
- The phylogeny of development and the origin of homology. In: Grande L & Rieppel O (eds) Interpreting the Hierarchy of Nature. Academic Press, San Diego.
- Stotz K (2005)
- Geschichte und Positionen der evolutionären Entwicklungsbiologie. In: Krohs U & Toepfer G (Hg) Philosophie der Biologie. Frankfurt, S. 338-356.
- Summerbell D, Lewis JH & Wolpert L (1973)
- Positional information in chick limb morphogenesis. Nature 244, 492-496.
- Tickle C (2006)
- Making digit patterns in the vertebrate limb. Nature Reviews. Molecular Cell Biology 7, 45-53.
- Ullrich H (1994)
- Embryologie und Homologie: Die Reichert-Gauppsche Theorie. Stud. Int. J. 1, 15-24.
- Ullrich H (2004)
- The „human tail“. Entwicklungsstörung oder Hinweis auf geschwänzte Vorfahren des Menschen? Stud. Int. J. 11, 51-58.
- Ullrich H (2005)
- Das Ende der klassischen Homologisierung? Neue Methoden in der vergleichenden embryonalen Forschung. Stud. Int. J. 12, 58-65.
- Vargas AO & Fallon JF (2005a)
- Birds have dinosaur wings: the molecular evidence. J. Exp. Zool. (Mol. Dev. Evol.) 304B, 85-89.
- Vargas AO & Fallon JF (2005b)
- The Digits of the Wing of Birds Are 1, 2, and 3. A Review. J .Exp. Zool. (Mol. Dev. Evol.) 304B, 206-219.
- Wagner GP & Gauthier JA (1999)
- 1, 2, 3 = 2, 3, 4: a solution to the problem of homology of the digits in the avian hand. Proc. Nat. Acad. Sci. USA 96, 5111-5116.
- Wagner GP (2005)
- The developmental evolution of avian digit homology: An update. Theor. Biosci. 124, 165-183.
- Walker A (1985)
- The braincase of Archaeopteryx. In Hecht MK et al. (eds) The beginning of birds. Proceedings of the International Archaeopteryx Conference Eichstätt, pp 123-134.
- Welten MCM et al. (2005)
- Gene expression and digit homology in the chicken embryo wing. Evol. Dev. 7, 18-28.
- Wray GA & Abouheif E (1998)
- When is homology not homology? Curr. Opin. Genet. Dev. 8, 675-680.
- Zhou Z, Barret PM & Hilton J (2003)
- An exceptionally preserved Lower Creataceous ecosystem. Nature 421, 807-814.
|
|  |