 |
Das Ende der klassischen Homologisierung?
Neue Methoden in der vergleichenden embryonalen Forschung
von Henrik Ullrich
Studium Integrale Journal
12. Jahrgang / Heft 2 - Oktober 2005
Seite 58 - 65
|
 |
Zusammenfassung: Zur Bedeutung der Embryologie für die Aufhellung stammesgeschichtlicher Zusammenhänge existieren spätestens seit den Veröffentlichungen von Haeckels vielzitierten und kritisierten Embryonentafeln und seinem Biogenetischen Grundgesetz sehr unterschiedliche Anschauungen. Insgesamt, so schätzen Richardson et al. ein, gelang bisher keine generelle Integration von ontogenetischen* Studien in die phylogenetische* Modellbildung aufgrund methodischer und terminologischer Schwächen der klassischen vergleichenden Embryologie sowie des hohen Komplexitätsgrades ontogenetischer Entwicklungsprozesse in Raum und Zeit. Neu entwickelte Methoden werden vorgestellt, die auf computergestützten Analysen quantifizierbarer embryonaler Daten beruhen: „event pairing“ und „3D-patterning“. Damit gewonnene Ergebnisse liefern eine objektivere Grundlage für vergleichende Studien und erlauben eine bessere Integration embryologischer Daten in phylogenetische Modelle im Vergleich zu den klassischen Vergleichsmethoden. Die neuen Methoden können allerdings keine Antwort auf Frage nach den ursächlichen Mechanismen hinter dem vorausgesetzten phylogenetischen Wandel geben. Die kausale Begründung der Dimension des phylogenetischen Wandels von Ontogenesen in der Zeit und nicht allein dessen hypothetische Ableitung – wie hier auf rein quantitativer Ebene –, bleibt die große Herausforderung für die Embryologie im Kontext der aktuellen „Evo-Devo“-Debatte.
|
|
|
Einleitung
Die Erforschung der Stammesgeschichte der Lebewesen verfolgt zwei Ziele: Zum einen sollen die Abstammungsverhältnisse rekonstruiert (Phylogenetik), zum anderen die Mechanismen des Formenwandel aufgeklärt werden (kausale Evolutionsforschung). In den vergangenen 150 Jahren beteiligte sich die Embryologie an der phylogenetischen* Forschung in beiden Sparten mit wechselndem Erfolg und Einfluß. Darwin sah in der Stammesgeschichte eine entscheidende Quelle für das Verständnis der Ontogenese*, jedoch nie in der extremen Form Haeckels. Das von Ernst Haeckel (1866,1872) propagierte Rekapitulationsdenken erwies sich als besonderer Kristallisationspunkt zahlreicher Studien und vehement geführter Auseinandersetzungen hinsichtlich der Genese embryonal auftretender Strukturen und ihrer homologen* Zuordnung zu adulten* oder embryonalen Merkmalen unterschiedlicher Organismen. Als bekannteste ontogenetische Bildungen dafür stehen die sogenannten „Kiemenbogen“, „Kiemenbogenarterien“, „Kiementaschen“ und „Kiemenspalten“, die in der Embryonalentwicklung nahezu aller Wirbeltiere, inklusive der des Menschen, auftreten und häufig als phylogenetische Reminiszenzen an fischartige Vorfahren zitiert werden. Ausgehend vom Paradigma Evolution suchte Haeckel mit dem Biogenetischen Grundgesetz „den engen, ursächlichen Zusammenhang…“ (Welträthsel 1903, S. 36) zwischen Phylogenese und Ontogenese darzustellen und gleichzeitig einen deskriptiv* faßbaren Zusammenhang als hilfreichen Leitfaden für die Stammbaumrekonstruktion festzuschreiben.
Das Biogenetische Grundgesetz, wie es Ernst Haeckel verstand, war bereits am Anfang des 20. Jahrhunderts wissenschaftlich widerlegt. Als eigenständige methodische Grundlage für die Phylogeneseforschung konnte es aufgrund der begrifflichen und inhaltlichen Unschärfe niemals dienen. Dem Namen nach überlebte es dennoch weiter im Bereich der vergleichenden Embryologie und Morphologie, wenn auch in stark eingeschränkter und modifizierter Form (z.B. als Biogenetische Grundregel). Zwei grundsätzliche Wege zeichneten sich dabei ab (vgl. Remane 1952, Sudhaus & Rehfeld 1992). Nur selten wird die Methode der direkten Übertragung von Organontogenesen auf den phylogenetischen Werdegang des Organs praktiziert. Wenn die Paläontologie oder die Systematik über den Homologievergleich* keine verwertbaren Ansätze liefern kann, kann dieser Ansatz helfen, hypothetische Übergangsstadien zu finden. Häufig zitierte Beispiele sind die Ableitung der Chordatiere aus den Larven sessiler (festsitzender) Manteltiere (Tunikaten; Romer & Parsons 1991) und die Reichert-Gauppsche Theorie zur Ableitung der Mittelohrknochen bei den Säugetieren (Ullrich 1994).
Der indirekte Weg, welcher der methodisch wichtigere ist und für viele Forscher als einzig möglicher gilt, orientiert sich an der üblichen Vorgehensweise der vergleichenden Homologieforschung. Alle während der Embryonalentwicklung bei einem Individuum auftretenden morphologischen Merkmale werden gleichberechtigt deskriptiv erfaßt. In einem zweiten Schritt erfolgt der Versuch der Homologisierung dieser Merkmale mit embryonalen oder adulten* Einzelmerkmalen anderer Individuen zur Klärung möglicher stammesgeschichtlicher Verwandtschaftsbeziehungen (z.B. Homologisierung der caudalen Knospe beim menschlichen Embryo mit dem Wirbeltierschwanz, Ullrich 2004).
Während Haeckel die Ontogenese durch die Phylogenese kausal zu begründen suchte, geht die aktuelle kausale Evolutionsforschung von einem entgegengesetzten Ansatz aus. Veränderungen ontogenetischer Prozesse werden danach als Ursache des phylogenetischen Wandels diskutiert. Garstang prägte dazu schon 1922 den richtungsweisenden Ansatz: „Ontogeny does not recapitulate phylogeny, it creates it“ (1922, 81).
Vor diesem Hintergrund wirkt eine aktuelle Einschätzung von Richardson & Verbeek (2003) wie eine Provokation: „Besitzt die klassische Form der vergleichenden Anatomie in der Embryologie eine Zukunft im Zeitalter der molekularen Biologie und der digitalen Technologien? Genauer, können in der vergleichenden Embryologie morphologische Merkmale weiterhin genutzt werden oder wird man neue Typen von Merkmalen definieren? … Auf den ersten Blick erscheint es so, daß in der vergleichenden Embryologie die traditionellen morphologischen Merkmale nicht länger benötigt werden“ (Richardson & Verbeek 2003, 303).
 |
Statt wie Haeckel, der die Ontogenese
durch die Phylogenese erklären wollte,
versucht die aktuelle kausale
Evolutionsforschung den umgekehrten
Erklärungsweg zu gehen. |
 |
|
Die Autoren hinterfragen damit die über ein Jahrhundert gepflegte und bislang vorherrschende Methodik des Merkmalsvergleichs in der vergleichenden Embryologie: Die Auswertung und den Vergleich deskriptiv erfaßter morphologischer Merkmale von Embryonen. Das würde nichts anderes als das Ende der klassischen Homologisierung bedeuten; man kann fast von der Aufgabe eines Paradigmas in der Evolutionsbiologie sprechen.
Die Kritik an der klassischen Homologisierung erfolgt aus mehreren Gründen. Richardson und Verbeek zeigen in der zitierten Arbeit die Grenzen der geschichtsträchtigen vergleichenden embryonalen Forschung für die Klärung phylogenetischer Fragestellungen auf. Die bisherigen, im einleitenden Abschnitt geschilderten Prozeduren des direkten und indirekten Weges haben sich nach ihrer Auffassung als wenig hilfreich erwiesen. Sie plädieren deshalb für einen anderen Ansatz, morphologische und genetische Befunde aus der Embryologie für stammesgeschichtliche Studien zu verwenden.
Die Autoren stellen neu entwickelte Methoden vor, die auf computergestützten Analysen quantifizierbarer embryonaler Daten beruhen, nämlich „event pairing“ und „3D-patterning“. Diese Werkzeuge sollen zum einen den Veränderungen des sich in Zeit und Raum entwickelnden Organismus besser gerecht werden. Damit gewonnene Ergebnisse würden zweitens eine objektivere Grundlage für vergleichende Studien liefern und deshalb eine bessere Integration embryologischer Fakten in den phylogenetischen Gesamtrahmen ermöglichen.
Nach einer kurzen Zusammenstellung der Hauptkritikpunkte der Arbeitsgruppe um Richardson an den klassischen Wegen der vergleichenden Embryologie sollen die vorgeschlagenen neuen Methoden etwas ausführlicher erläutert werden. Ob damit die angesprochenen Probleme der Integration ontogenetischer Fakten in phylogenetische Zusammenhänge besser gelöst werden, wird abschließend diskutiert.
|
adult: erwachsen, ausgereift, voll entwickelt. Allometrisches Wachstum: disproportionales Wachstum von Organen/Strukturen, bei welchem Verschiebungen von Relationen einzelner Organe zueinander auftreten; bekannter mikroevolutiver Prozeß (z.B. bei Geweihen, Zähnen, Hörnern). deskriptiv: beschreibend. Entwicklungszwänge (constraints): genetisch und epigenetisch (z.B. morphologisch, physiologisch) bedingte Einschränkungen, die einer beliebigen phylogenetischen Abwandlung embryonaler Abläufe entgegenstehen. Expression, Genexpression: Prozeß der Übersetzung genetischer Informationen in Eiweißverbindungen. Zu einem bestimmten Zeitpunkt werden immer nur bestimmte Gene des Genoms exprimiert. Heterochronie: beim Vergleich von Entwicklungssequenzen verschiedener Tierarten nachweisbare Unterschiede hinsichtlich der Reihenfolge des Erscheinens bzw. der Dauer des Nachweises von embryonalen oder ontogenetischen Merkmalen (z.B. der Herzanlage bei Säugetieren und Reptilien) Homologie: Ähnlichkeit von Strukturen, Gensequenzen, Biomolekülen usw. bei Organismen, die im phylogenetischen Kontext auf eine gemeinsame stammesgeschichtliche Herkunft zurückgeführt wird. Konvergenz: Ähnlichkeit von Strukturen, Gensequenzen, Biomolekülen u.s.w. bei Organismen, die im phylogenetischen Kontext nicht auf eine gemeinsame stammesgeschichtliche Herkunft zurückgeführt wird. Ontogenese: Individualentwicklung von Organismen vom Zeitpunkt der Befruchtung bis zur Geburt bzw. dem Schlüpfen aus dem Ei; unterteilt in Blastogenese, Embryogenese, Fetogenese. Phylogenese: Theorie der stammesgeschichtlichen Herkunft aller heutiger Lebensformen durch kontinuierliche Abwandlung von Vorgängerarten auf der Grundlage natürlicher Prozesse.
|
|
|
Probleme bei der Integration ontogenetischer Studien in phylogenetischen Rekonstruktionen
Wie bereits am Anfang erwähnt, existieren zur Bedeutung der Embryologie für die Aufhellung stammesgeschichtlicher Zusammenhänge spätestens seit den Veröffentlichungen von Haeckels viel zitierten und kritisierten Embryonentafeln und seinem Biogenetischen Grundgesetz sehr unterschiedliche Anschauungen. Insgesamt, so schätzen Richardson et al. (2001) ein, gelang bisher keine generelle Integration von ontogenetischen Studien in die phylogenetische Methodologie und Modellbildung. Ursachen sind nach ihrer Ansicht auf drei unterschiedlichen Ebenen auszumachen.
| Abb. 1: Portrait-Galerie und Sanduhrmodell. Die klassische Portrait-Galerie der vergleichenden Embryologie (links, online hier leider nicht verfügbar) und das daraus abgeleitete Sanduhrmodell (rechts). Der engste Punkt des Sanduhrmodells entspricht der obersten Reihe der Embryonengalerie und dem so genannten „phylotypischen Stadium“. Beide Schemata repräsentieren die Ansicht, daß ein bestimmtes Stadium der embryonalen Entwicklung bei allen Wirbeltieren die höchste Ähnlichkeit untereinander aufweist, während der Beginn (Frühentwicklung) und die späteren Phasen der Entwicklung (embryonale und fetale Spezialisierung) größere Divergenzen aufweisen. Das Sanduhrmodell wurde in breit angelegten Studien widerlegt. (Links nach Richardson 1998, rechts nach Richardson 1999) |
 |
|
1. Zunächst entwickelten sich die modernen phylogenetischen Methoden der Synthetischen Evolutionstheorie, nachdem die Embryologie am Anfang des 20. Jahrhunderts ihre zentrale Stellung in der Evolutionsbiologie verloren hatte (Gould 1977). In dieser Zeit verschob sich der Schwerpunkt der embryonalen Forschung zur experimentellen Seite, die Suche nach universellen ontogenetischen Entwicklungsmechanismen oder -prinzipen stand im Vordergrund. Nicht mehr eine breite Anzahl unterschiedlicher Arten, die in den so genannten „Portrait-Galerien“ einander gegenübergestellt und entsprechend phylogenetisch ausgewertet wurden, stand im Zentrum des Interesses (vgl. Abb. 1). Die Experimentalisten wählten vielmehr Modellorganismen, die im Labor gut zu untersuchen waren (z.B. Fruchtfliege, Fadenwurm, Maus, Zebrafisch, Seeigel) und konnten dabei bis heute große Erfolge im Verständnis der Entwicklungsvorgänge auf molekulargenetischer Ebene, speziell der Musterbildung, der räumlichen und zeitlichen Differenzierung (z.B. durch Hox-Gene) gewinnen. Weniger erfolgreich waren dagegen die Versuche, diese Daten in eine neue Synthese von Evolution und Entwicklung („Evo-Devo“) zu integrieren.
2. Die phylogenetisch arbeitenden Entwicklungsbiologen konzentrierten sich beim anatomischen Vergleich von Ontogenesen zumeist auf „konservierte“ Merkmale, die für sie im Sinne Haeckels als repräsentativ für adulte oder embryonale Strukturen stammesgeschichtlicher Vorfahren galten (z.B. die embryonalen Pharyngealbögen bei den Wirbeltieren als Rekapitulation adulter bzw. embryonaler Kiemenanlagen). Niemals konnte jedoch eine widerspruchsfreie Methode und Terminologie für diese Form des Vergleichs gefunden werden. Zentrale Fragen konnten nicht befriedigend im Sinne phylogenetischer Zusammenhänge gelöst werden:
- Welche Merkmale können in den Vergleich einbezogen werden und welche nicht? (d. h.: Welche Merkmale geben in phylogenetischer Deutung einen Hinweis auf stammesgeschichtliche Vorfahren?)
- Welche sind repräsentativ für eine Homologisierung über die Artgrenzen hinaus und in Bezug auf welches adulte, ausgereifte Organ? (Vgl. dazu Abb. 2!).
- Ist das Fehlen eines embryonalen Merkmals beim Vergleich von Organismen als primitiv oder als sekundär verloren anzusehen?
- Gibt es eine Phase, in der Embryonen der Wirbeltiere eine größere phylogenetisch bedingte Ähnlichkeit aufweisen als in anderen?
 |
| Abb. 2: Homologieproblematik am Beispiel der Entwicklung von Pulmonal- und Aortenklappen. Ein typisches Homologieproblem der Entwicklungsbiologie, welches bei Nutzung gelisteter anatomischer Termini entsteht. A, B: schematische Querschnitte des Ausflußtraktes des frühen embryonalen Herzens beim Huhn (A) und beim Menschen (B). Das Huhn zeigt zwei vordere Wülste (dunkel- und hellblau) der Mensch nur einen (dunkelblau). C und D repräsentieren die Situation, nachdem der Ausflußtrakt separiert ist (Aorta, Pulmonalarterie) und die Gefäßklappen sich gebildet haben. Diese zeigen äußerlich keine wesentlichen Unterschiede, obwohl die embryonalen Ausgangsstrukturen z. T. unterschiedlich sind. Da die anatomischen Strukturen und deren Bezeichnungen in A und B nicht identisch sind, bleiben die Homologien zwischen C und D unklar (aus Richardson 2003). |
|
Gerade die zuletzt aufgeworfene Problematik sorgt aktuell für viel Diskussion. Haeckel behauptete, daß ein „Pharyngulastadium“ von allen Embryonen der Wirbeltiere durchlaufen wird. Dieses Stadium besäße die höchste Relevanz für die vergleichende Forschung, weil in diesem Stadium keinerlei wesentliche Unterschiede zwischen den Embryonen nachweisbar wären. Diese Behauptung hat sich schnell als falsch erwiesen (vgl. Ullrich 1998). Dennoch wurde an der Idee festgehalten, Stadien zu definieren, denen ein höherer phylogenetischer Wert zukommen soll gegenüber anderen ontogenetischen Entwicklungsstadien. Die Entwürfe zur „Körpergrundgestalt“ (Seidel 1960), eines phyletischen (Cohen 1979) oder des phylotypischen (Sander 1983) Stadiums oder des Phylotypus (Slack et al. 1993) und des Zootypus (Hall 1992) sind Ausdruck dieses Bemühens, beobachtbare Ähnlichkeiten zwischen den Embryonen in verschiedenen Entwicklungsphasen mit einer besonderen phylogenetischen Bedeutung zu belegen.
Es fehlt jedoch ein objektivierbares methodisches Kriterium, um mit Sicherheit ein Stadium oder ein Merkmal gegenüber anderen als „universeller“ oder „konservierter“ und damit für die Rekonstruktion der Phylogenese besonders relevant zu kennzeichnen. Daher ist es unmöglich, sichere bzw. objektivierbare Homologisierungen festzulegen, die als Wegweiser für die Rekonstruktion der Stammesgeschichte dienen könnten. Für die phylogenetische Auswertung erweisen sich damit keine Merkmale oder Stadien als aussagekräftiger als andere.
Außerdem konnten Richardson et al. durch vergleichende Studien zeigen, daß kein phylogenetisch konservatives Stadium bei Wirbeltieren existiert. Der Grad an Ähnlichkeiten in den einzelnen Entwicklungsphasen unterschiedlicher Wirbeltierarten stellt sich entgegen dem „Sanduhrmodell“ auch in dem als „konservativ“ bezeichneten Stadium nicht größer als in anderen dar (Richardson 1995,1997, siehe dazu Abb. 1).
3. Als drittes Problem verweisen die Autoren auf die besondere Natur von Entwicklungsprozessen und -merkmalen. Die Ontogenese muß als ein vierdimensionaler Prozeß analysiert werden, drei Dimensionen betreffen die Veränderungen der Einzelmerkmale im Raum (Länge, Breite, Tiefe) und die vierte deren Wandel in der Zeit. Die phylogenetische Analyse kommt als fünfte Dimension dazu, da hier die Unterschiede und Gemeinsamkeiten von Entwicklungswegen über die Artgrenzen hinaus vergleichend betrachtet werden. Um den dargestellten Wandel ontogenetischer Merkmalskonstellationen in Raum und Zeit abbilden zu können, reicht nach Richardson et al. eine rein deskriptive Terminologie nicht mehr aus. Identische Bezeichnungen wie „Herzschlauch“, „Wülste“, „Gefäßklappen“, „Aorta“, „Pulmonalarterie“ usw. werden für entsprechende ontogenetische Einzelmerkmale in unterschiedlichen Entwicklungsphasen benutzt, obwohl das betrachtete Merkmal einen drastischen Form- und Größenwandel durchläuft. Gerade solche wesentlichen Änderungen können auf der Grundlage der bislang genutzten, deskriptiv orientierten Terminologie weder adäquat widergespiegelt noch analytisch ausgewertet werden.
Diese terminologische Unschärfe im Zusammenhang mit der ontogenetischen Komplexität scheint zunächst undurchdringbar. Richardson et al. hoffen jedoch, sie dank neuer Möglichkeiten der vergleichenden Forschung beherrschbar zu machen. Darauf kommen wir im folgenden zu sprechen.
|
|
Event pairing und 3D-patterning – Quantifizierung entwicklungsbiologischer Daten im phylogenetischen Kontext
Der methodische Ausweg aus den geschilderten Problemen der traditionellen vergleichenden Entwicklungsbiologie, die sich meist auf den „nicht quantitativen“ Vergleich von „Portrait-Galerien“ stützte, wird von Richardson und seiner Arbeitsgruppe in neuen computergestützten Analysen quantifizierbarer embryonaler Daten gesehen. Dabei werden die in der Ontogenese von Individuen auftretenden Merkmale unter Berücksichtigung ihrer räumlichen bzw. zeitlichen Charakteristik auf molekular-genetischer oder anatomischer Ebene quantitativ erfaßt. In einem zweiten Schritt erfolgt der Vergleich dieser Daten mittels Computeranalysen innerhalb und außerhalb der Artgrenzen als Grundlage für eine phylogenetische Interpretation.
Event pairing
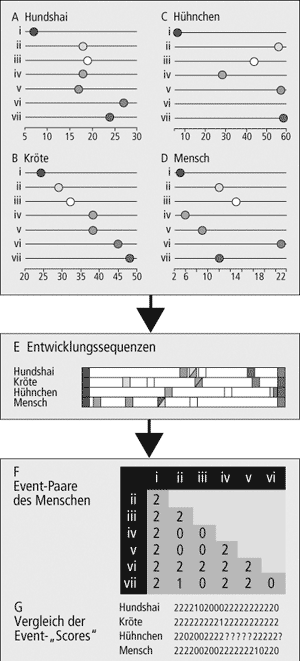 |
Abb. 3: Methodik des „Event pairing“
A-D Das erste individuelle Auftreten von sieben entsprechenden embryonalen Strukturen (i: erster Somit, ii: Nasenplacode, iii: Augenbecher, iv: Herzschlauch, v: Schilddrüsenanlage, vi: Milzanlage, vii: Knospen der vorderen Extremitäten) bei vier Wirbeltierarten (A Hundshai, B Kröte, C Hühnchen, D Mensch) wird durch einen farbigen Punkt markiert (= „event“). Die x-Achse repräsentiert die Entwicklungszeit und ist in Entwicklungsstadien untergliedert. Die Entwicklungsstadien für jede Art können jedoch nicht direkt miteinander verglichen werden. Angaben zur Milzentwicklung beim Hühnchen fehlen.
E Entwicklungssequenzen: Die lineare Anordnung des ersten Auftretens der o.g. embryonalen Strukturen führt zu Entwicklungssequenzen, die sich artübergreifend vergleichen lassen. Die dabei faßbaren Differenzen zeigen jedoch keine absoluten sondern nur relative Unterschiede bezüglich des Zeitpunktes des Auftretens dieser Strukturen auf.
F Event-Paare: Darstellung einer „event pair-matrix“ für den Menschen. Dabei wird jedes Merkmal (i-vii) in Relation zu jeweils einem anderen gesetzt und bewertet. Der Wert („Score“) „0“ steht dafür, daß das Merkmal vor, bei „1“ zeitgleich mit und bei „2“ nach dem anderen Merkmal auftritt.
G Scorevergleich: Im Ergebnis lassen sich in einer definierten Reihenfolge (von i nach vii) die 21 möglichen Merkmalsverhältnisse als Zahlenwerte formulieren (z.B.: Mensch: 222200200222222210220) und mit denen anderer Tierarten (B-D) vergleichen und statistisch auswerten. (Nach Jeffery et al. 2002b) |
|
Der Entwicklungsweg eines Individuums wird in eine Serie von Einzelereignissen („events“) untergliedert, wobei diese als Änderungen von Zuständen (Auftreten oder Verschwinden eines Merkmals, z. B. der Augenanlage, der Herzanlage, der Extremitätenknospen u.s.w.) definiert werden (vgl. dazu Abb. 3). Die einzelnen Ereignisse dokumentieren gleichzeitig den Beginn oder das Ende der Expressionsaktivität* eines Gens oder eines Genclusters. Eine Serie dieser Ereignisse, geordnet in chronologischer Reihenfolge, wird als Entwicklungssequenz bezeichnet.
Die Schlüsselmethode zur quantitativen Analyse einer solchen Sequenz ist das „event pairing“, die paarweise Erfassung aller Ereignisse einer Sequenz und damit der relativen zeitlichen Beziehungen aller Ereignisse zueinander. Zum Beispiel mag in einer Art das Ereignis „X“ vor dem Ereignis „Y“ auftreten. Im Vergleich mit Sequenzen anderer Arten kann nun geklärt werden, ob das Ereignis „Y“ vor „X“, gleichzeitig mit „X“ oder nach „X“ auftritt. Die Unterschiede der „event pairing“-Datensätze zwischen verschiedenen Arten sind Ausdruck einer anderen zeitlichen Steuerung (Timing) von Entwicklungsereignissen hinsichtlich ihres Beginns oder ihrer Dauer in den jeweiligen individuellen Entwicklungssequenzen. Unter dem Begriff „Heterochronien“* sind diese zwar schon seit Haeckel bekannt und vielfach diskutiert, können nun aber mit einem Zahlenwert („Score“) von 0 bis 2 (0: „X“ tritt vor „Y“ auf; 1: „X“ tritt mit „Y“ auf , 2: „X“ tritt nach „Y“ auf) quantifiziert werden. Bei einer großen Anzahl von betrachteten Ereignissen lassen sich dann diese Ergebnisse nutzen, um statistisch vergleichende Analysen durchzuführen und phylogenetische Fragestellungen anzugehen (z.B. bei Jeffery et al. 2002, Binanda-Emonds et al. 2003).
Eine Modifikation dieser Technik, das „event pair cracking“, erlaubt Schlüsselheterochronien* zu bestimmen, denen dann besonderes Gewicht bei der Klärung der Diversifikation innerhalb phylogenetischer Linien zukommt. Dabei werden Einzelereignisse gegenüber einem Block definierter „event pairs“ (das sind meist als abgeleitete bzw. apomorph eingestufte Merkmale) getestet, um zeitliche Verschiebungen von Einzelereignissen bei unterschiedlichen Arten als aktiven oder passiven (d.h. mitgeschleppten) Vorgang zu erfassen (Mabee & Trendler 1996, Smith 1996).
Zusätzlich ist es möglich, mit dem „event pairing“ Veränderungen des Entwicklungsablaufes bei unterschiedlichen Rahmenbedingungen (z.B. Sauerstoffgehalt bei der Entwicklung des cardiovaskulären Systems von Knochenfischen) innerhalb einer Art zu studieren. Dadurch erhofft man sich einen quantitativen Zugang, um die oftmals erstaunliche innerartliche phänotypische Plastizität besser zu charakterisieren.
3D-patterning
 |
Abb. 4: 3D-patterning.
Ein voxelbasierter Ansatz zum Vergleich von räumlichen Mustern der Genexpression. Oben wird ein Arrangement von Volumenelementen (Voxel) gezeigt, jedes besitzt eine eigene Nummer (sichtbar an der Oberfläche der Voxel), jedes Voxel repräsentiert einen Charakter, welcher in vergleichenden Untersuchungen genutzt werden kann. Der Status („A“, „B“, „AB“) jedes Charakters wird in diesem Schema vom Muster der Genexpression zweier Gene („A“, „B“) bestimmt, wie unten gezeigt. Schwarz: Expressionsdomäne von Gen A, hellblau von Gen B, dunkelblau: von beiden Genen. (Nach Richardson 2003) |
|
Schon lange sind Techniken zur Rekonstruktion dreidimensionaler Modelle anatomischer Strukturen während der Embryonalentwicklung bekannt (z. B. Blechschmidts Rekonstruktion der menschlichen Ontogenese) und in vielfacher Anwendung. Neu ist nun der Versuch, diese räumlichen Modelle in eine computergestütze Datenbank zu übertragen und diese wiederum über die Artgrenzen hinaus zu vergleichen. Entscheidend dabei ist die Festlegung und Definition einer Matrix als Referenzsystem der zu vergleichenden räumlichen und zeitlichen Merkmalsmuster. Diese Matrix existiert bereits für einzelne Modellobjekte der Embryologie, nämlich von der Maus und dem Zebrafisch (Baldock et al. 1997, Verbeek et al. 1998, 2002). Als Daten werden histologische (Gewebs-) Merkmale erfasst, die einerseits einem anatomischen Kennzeichen, anderseits exprimierten Genen oder Genclustern zugeordnet sind. Letztere repräsentieren gleichzeitig eine Art Status des anatomischen Merkmals in der Zeit (vgl. Abb. 4). Um den Unschärfen bei der Verwendung anatomischer Begriffe zu begegnen, die ja im Verlauf der Entwicklung keine Konstanten innerhalb einer Art und erst recht über die Artgrenzen hinaus darstellen, bemüht man sich, die Matrix durch Volumenelemente (sog. „Voxel“) und den Status der anatomischen Kennzeichen als Liste exprimierter Gene zu definieren. Bisherige anatomische Bezeichnungen (wie z.B. „chorda dorsalis“ oder „Entoderm“) dienen dann „nur“ noch als Referenzen bei der Etablierung solcher Systeme im Zusammenhang mit der ersten Datenaufnahme oder als Orientierungshilfe bei der Untersuchung des entsprechenden embryonalen Objekts.
Das Voxel kann eine Zelle, einen Zellverband oder ein Gewebe repräsentieren. Der Vergleich dieser voxelbasierten Datensätze über die Artgrenzen hinaus erlaubt dann wiederum quantitative Aussagen zu Ähnlichkeiten oder Unterschiede auf der Ebene raum-zeitlicher Differenzierungen.
|
|
Neue Methoden und alte Probleme
Richardson ist zuzustimmen, daß neue, tiefere und sicher auch faszinierende Einsichten auf der Grundlage der oben geschilderten Methoden für den Vergleich der komplexen, sich in vier Dimensionen vollziehenden Entwicklungsverläufe von Organismen zu erwarten sind. Die Kapazitäten heutiger Computer lassen es zu, die zahllosen Daten verschiedener Arten unter mehreren Aspekten auszuwerten (innerartlich bzw. überartlich, organspezifisch, funktionsspezifisch u.s.w.). Bei aller Euphorie bleibt jedoch noch eine Fülle an Detailarbeit zu tun. Die Erforschung und exakte Beschreibung der zahllosen Ontogenesen von Tieren unterschiedlicher Stämme ist erforderlich, um eine entsprechend breite Datenbasis für die neuen Methoden zur Verfügung zu stellen.
Die bisher vorliegenden Ergebnisse bei Nutzung des „event pairing“ haben bereits manch lieb gewonnene Vorstellung, die auf der Basis der herkömmlichen Methoden erstellt wurde, in Frage gestellt. Nachdem Richardson bereits mit dem klassischen Vorgehen nachwies, daß kein „phylotypisches Stadium“ bei den Wirbeltieren existiert (1995, 1997), belegten Bininda-Emonds et al. (2003a) dies quantitativ. Sie zeigten, daß gerade in der dafür favorisierten mittleren embryonalen Phase die morphologischen Differenzen und die Verschiebungen in den entsprechenden Entwicklungssequenzen bei Wirbeltieren (Heterochronien) am stärksten ausgeprägt sind. Die Idee von einem besonders konservierten und daher phylogenetisch signifikanten Embryonalstadium kann damit wohl als widerlegt gelten.
|
Die Idee von einem besonders
konservierten und daher phylogenetisch
signifikanten Embryonalstadium
kann wohl als widerlegt gelten. |
|
|
Eine andere Untersuchung beschäftigte sich mit einem bislang favorisierten Mechanismus an der Wurzel der Säugetierphylogenese. Mit der Methode des „event pairing“ ging man der Fragestellung nach, ob durch Heterochronien deren große Diversifikation zu erklären ist. Gould (1977, 1982), Raff (1996) und andere vermuteten, daß insbesondere Veränderungen von Wachstumsraten (allometrische* Heterochronien) in frühen embryonalen Phasen hinter dem evolutionären Wandel der Säugetiere stehen könnten. Die Ergebnisse von Bininda-Emonds et al. (2003b) konnten dies nicht bestätigen. Die 116 untersuchten embryonalen Entwicklungsereignisse („events“) zeigen erstaunlicherweise zwischen den Säugetiergruppen (z.B. Paarhufer, Primaten, Nagetiere) kaum Unterschiede hinsichtlich ihres relativen zeitlichen Auftretens beim Vergleich der jeweiligen Entwicklungssequenzen. Beim Vergleich mit anderen Gruppen der Wirbeltiere (vor allem Tetrapoda) sind demgegenüber ausgeprägte Heterochronien (s.o.) nachweisbar. Die Autoren schlußfolgern, daß spezielle Entwicklungszwänge* („constraints“) am Beginn der Säugetierevolution bestanden, die einem evolutionären Wandel der Entwicklungssequenzen in frühen embryonalen Stadien durch Heterochronie entgegenstanden. Welche Mechanismen zur Radiation der Säugetiere führten, bleibt weiter offen. In einzelnen Fällen werden postembryonal wirksame Heterochronien (z.B. durch allometrisches Wachstum* der Flosse beim Vorfahren von Delphinen) favorisiert.
Die Auswertung der quantifizierten embryonalen Daten wird auch für die Erstellung von „Stammbäumen“ genutzt. Die ersten Ergebnisse für eine Gruppe von Wirbeltieren (Amnioten) entsprechen fast vollständig den vorliegenden cladistischen phylogenetischen Systemen (Jeffery et al. 2002). Nur die Einordnung der braunen Ratte korreliert nicht mit dem bisherigen Schema und wirft entsprechende Fragen hinsichtlich der phylogenetischen Einordnung der Nagetiere auf.
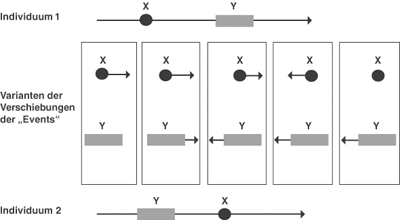 |
| Abb. 5: Sequenzverschiebung und fünf mögliche Ursachen. Das embryonale Merkmal „X“ tritt beim Individuum 1 vor dem Merkmal „Y“ auf, beim stammesgeschichtlich abgeleiteten Individuum 2 ist die Reihenfolge umgekehrt. Es gibt fünf Varianten, wie es zu dieser Verschiebung theoretisch kommen kann. Die Verschiebungen der „Events“ werden durch Pfeile gekennzeichnet, deren Länge Ausdruck des relativen Ausmaßes des Versatzes innerhalb der Entwicklungssequenz ist. Mit „event pairing“ kann nur die unterschiedliche Reihenfolge der „Events“ beschrieben, jedoch keine Variante als tatsächlicher Weg favorisiert werden. |
|
Die neuen Werkzeuge sind aber nicht frei von methodischen Schwachpunkten und können auch nicht alle alten inhaltlichen Probleme der vergleichende Forschung lösen. Ein Nachteil der „event pairing“-Methode wird darin gesehen, daß nur relative Unterschiede zwischen Ereignissen von Entwicklungssequenzen angezeigt werden können. Treten zum Beispiel die Merkmale „X“ und „Y“ bei einer Art nacheinander und in einer zweiten in der umgekehrten Reihenfolge auf, kann nur der relative Versatz beider Ereignisse beschrieben werden. Wir wissen aber nichts darüber, welches dieser Ereignisse sich in welche Richtung, wann und warum es sich verschoben hat. Theoretisch lassen sich dafür fünf Verschiebungsmöglichkeiten ableiten, die fünf möglichen phylogenetischen Wegen entsprechen, ohne daß einer von der Methode her favorisiert werden kann (vgl. Abb. 5).
Ein zweites Problem besteht darin, daß Merkmalskombinationen, die phylogenetisch einander nicht beeinflussen sollten (z.B. Extremitätenknospen und Augenanlage) gleichberechtigt mit phylogenetisch einander bedingenden Merkmalskombinationen (Gehirnanlage mit Augenanlage) betrachtet und analysiert werden.
Ein drittes Kernproblem hat die neue Methode jedoch von den klassischen Methoden übernommen. Das deskriptive Feststellen von Unterschieden im Auftreten von Ereignissen in Entwicklungssequenzen und deren numerische Quantifizierung mit einem Zahlenwert (0; 1; 2) belegt und beweist nicht, daß es diese Verschiebungen in der Geschichte auch tatsächlich gegeben hat. „Event pairing“ vermag keine Antwort auf Frage nach den kausalen Mechanismen eines solchen Wandels im Raum-Zeit-Kontinuum des sich entwickelnden Organismus zu geben. Daran ändert sich auch nichts, wenn wir genau wissen, welche Genaktivitäten mit welchen morphologischen Merkmalen und ihren Änderungen gekoppelt sind (3D-patterning). Interessant wird sein, wie man argumentativ mit den „quantifizierbaren“ Konvergenzen* in Entwicklungssequenzen von Arten unterschiedlicher phylogenetischer Linien umgeht.
Die kausale Begründung der „5.“ Dimension, des phylogenetischen Wandels von Ontogenesen in der Zeit, und nicht allein dessen deskriptive hypothetische Darstellung auch auf rein quantitativer Ebene, bleibt die große Herausforderung für die Embryologie im Kontext der aktuellen „Evo-Devo“-Debatte.
Für die Schöpfungsforschung können die diskutierten methodischen Ansätze hilfreich sein, um andere Facetten des Variationspotentials innerhalb bekannter Grundtypen zu erfassen.
|
Literatur
- Baldock RA, Verbeek FJ & Vonesh JL (1997)
- 3D reconstructions for graphical databases of gene-expression. Semin. Cell Dev. Biol. 8, 499-507.
- Blechschmidt E (1961)
- Die vorgeburtlichen Entwicklungsstadien des Menschen. Freiburg.
- Bininda-Emonds ORP, Jeffrey JE & Richardson MK (2003a)
- Inverting the hourglass: quantitative evidence against the phylotypic stage in vertebrate development Proc. R. Soc. Lond. B 270, 341-346.
- Bininda-Emonds ORP, Jeffrey & JE Richardson MK (2003b)
- Is Sequence Heterochrony an Important Evolutionary Mechanism in Mammals? J. Mammal. Evol. 10, 335- 361.
- Garstang W (1922)
- The theory of recapitulation: a critical restatement of the biogenetic law. Zool. J. Linn. Soc. 35, 81-101.
- Gould SJ (1977)
- Ontogeny and Phylogeny. Belknap Press, Cambridge, Ma.
- Gould SJ (1982)
- Change in developmental timing as a mechanism of macroevolution. In: Bonner JT (ed) Evolution and Development. New York, pp. 333-346.
- Hall BK (1996)
- Baupläne, Phylotypic Stages and Constraint. Evol. Biol. 29, 215-261.
- Haeckel E (1866)
- Generelle Morphologie. I: Allgemeine Anatomie der Organismen. II: Allgemeine Entwickelungsgeschichte der Organismen. Berlin.
- Jeffery JE, Richardson MK, Coates MI & Bininda-Emonds ORP (2002a)
- Analyzing developmental sequences within a phylogenetic framework. Syst. Biol. 51, 478-491.
- Jeffery JE, Richardson MK, Coates MI & Bininda-Emonds ORP (2002b)
- Analysing evolutionary patterns in vertebrate embryonic development. Evol. Develop. 4, 292-302.
- Raff RA (1996).
- The Shape of Life: Genes, Development and the Evolution of Animal Form. Chicago.
- Remane A (1952)
- Die Grundlagen des natürlichen Systems Band 1. Leipzig.
- Richardson MK (1995)
- Heterochrony and the phylotypic period. Dev. Biol. 172, 412-421.
- Richardson MK, Hanken J, Gooneratne ML, Pieau C, Raynaud A, Selwood L & Wright GM (1997)
- There is no highly conserved embryonic stage in the vertebrates: implications for current theories of evolution and development. Anat. Embryol. 196, 91-106.
- Richardson MK, Jeffery JE, Coates MI & Bininda-Emonds, ORP (2001)
- Comparative methods in developmental biology. Zoology (Jena) 104, 278-283.
- Richardson MK & Keuck G (2002)
- Haeckel’s ABC of evolution and development. Biol. Rev. 77, 495-528.
- Richardson MK & Verbeek FJ (2003)
- New directions in comparative embryology and the nature of developmental characters. Animal Biology 53, 303-311.
- Sudhaus W & Rehfeld K (1992)
- Einführung in die Phylogenetik und Systematik. Stuttgart, Jena, New York.
- Ullrich H (1994)
- Embryologie und Homologie – Die Reichert-Gauppsche Theorie. Stud. Int. J. 1, 15-24.
- Ullrich H (1998)
- Die Wiederentdeckung eines Irrtums. Stud. Int. J. 5, 3-6.
- Ullrich H (2004)
- The „human tail“. Entwicklungsstörung oder Hinweis auf geschwänzte Vorfahren des Menschen? Stud. Int. J. 11, 51-58.
- Verbeek FJ & Huijsmans DP (1998)
- A graphical database for 3D reconstruction supporting different geometrical represantations. In: Wong STC (ed) Medical Image Databasis. Kluwer Academic Publishers Boston, pp117-144.
- Verbeek FJ, Boon PJ, Sloetjes H, van der Velde R & de Vos, N (2002)
- Visualization of complex data sets over Internet: 2D and 3D visualization of the 3D digital atlas of zebra sh development. Proc. SPIE (The International Society of Optical Engineering), 4672, 20-29.
|
|  |
