 |
The „human tail“
Entwicklungsstörung oder Hinweis
auf geschwänzte Vorfahren des Menschen?
von Henrik Ullrich
Studium Integrale Journal
11. Jahrgang / Heft 2 - Oktober 2004
Seite 51 - 58
|
 |
Zusammenfassung: Fehlbildungen der menschlichen Individualentwicklung der unteren Rückenanteile und des Steißbeins können sich in Form „schwanzartiger“ Anhängsel oder als Vorwölbungen über der Steißbeinspitze darstellen. Diese Strukturen werden häufig als prominentes Argument zitiert, um die Existenz von Atavismen* beim Menschen zu belegen – Rückschläge in stammesgeschichtlich frühere tierische Stadien. Die normalen anatomischen und ontogenetischen* Zusammenhänge der Entwicklung dieser Region werden den unterschiedlichen teratologischen (Mißbildungs-) Mustern und ihren möglichen Ursachen gegenübergestellt. Zusammen mit vergleichend anatomischen Betrachtungen sprechen die vorliegenden Befunde gegen eine Interpretation dieser Fehlbildungen als reaktiviertes Merkmal geschwänzter Vorfahren.
Ebenso ungerechtfertigt ist eine homologe* Gleichsetzung der embryonal auftretenden kaudalen* Knospe beim Menschen mit den echten Schwänzen von Wirbeltieren.
(* Mit einem Stern versehene Begriffe finden sich im Glossar am Ende des Kapitels „Geschichtlicher Rückblick“)
| |
Die Phänomene und ihre Darstellung
| Abb. 1: „True Tails“ Variante A: wurmartige Anhängsel in der Region über der Lendenwirbelsäule dem Kreuz- und Steißbein. |
|
Seit mehr als 150 Jahren werden in der wissenschaftlichen Literatur spezielle Fehlbildungen der menschlichen Individualentwicklung im Bereich der lumbosakralen Region (Übergang des Rückens von der Lendenwirbelsäule zum Kreuzbein) und des Steißbeins mit dem Begriff „human tail“ („menschlicher Schwanz“) belegt. Auch eine während der normalen embryonalen Entwicklung bei Wirbeltieren auftretende Struktur – als „caudal bud“, „Schwanzknospe“ oder „kaudale Eminenz“ bezeichnet – bringt man oft in Verbindung mit echten Schwänzen und diskutiert ihre Relevanz für stammesgeschichtliche Fragestellungen. Beides also, Resultate embryo-fetaler Entwicklungsstörungen bzw. teratologische* Strukturen und ontogenetisch* vorübergehend auftretende Bildungen gelten in diesem Interpretationsrahmen als Atavismen* bzw. als rekapitulierte* Merkmale geschwänzter Vorfahren des Menschen (Kunimoto 1918, Galiza 2001, Islam 2002).Im Folgenden sollen die oberdevonischen Fossilfunde vorgestellt und ihre Bedeutung für den Übergang Fisch – Vierbeiner diskutiert werden (s. Abb. 4). Demnach folgt auf Eusthenopteron der tetrapodenähnlichste Fisch Panderichthys und die ähnliche, aber nicht so gut erhaltene Gattung Elpistostege. Im Frasnium (unteres Oberdevon) sind Elginerpeton und Obruchevichthys überliefert, die als älteste Tetrapoden gelten. Allerdings weisen diese Gattungen einige deutlich spezialisierte Merkmale auf und werden deshalb auf einen blind endenden Seitenzweig gestellt. Eine interessante Gattung aus dieser Zeit ist außerdem Livoniana.
Eine einheitliche Klassifikation und Darstellung der angesprochenen Phänomene erwies sich wegen der verwirrenden Vielfalt bekannter Merkmalskombinationen als unmöglich. Für unsere Betrachtung relevant sind die als „true tails“ („wahre, echte Schwänze“) deklarierten Strukturen. Welche Fehlbildungen diesen Namen verdienen oder nicht, wird von den Autoren kontrovers diskutiert. In der Literatur wurden in den letzten 120 Jahren damit zumeist drei unterschiedliche Befundkomplexe beschrieben, zwei davon entsprechen echten embryonal-fetalen Fehlbildungen, die dritte einem normalen embryonalen Merkmalsgefüge. Davon abgegrenzt sind unter der Bezeichnung „Pseudotails“* meist äußerlich ähnliche Mißbildungen, die klinisch in Kombination mit bösartigen Tumorbildungen oder schweren Entwicklungsstörungen der Wirbelsäule und/oder des Zentralen Nervensystems auftreten. „Pseudotails“ besitzen für phylogenetische Betrachtungen keine wesentliche Bedeutung und werden an dieser Stelle nicht weiter berücksichtigt. Eine scharfe Abgrenzung der „Pseudotails“ gegen einzelne zu den „true tails“ gerechnete Strukturen ist jedoch nicht immer sicher möglich (Dubrow 1988, Dao 1983).
Wie erwähnt werden in der Literatur unter den „true tails“ meist zwei Gruppen embryo-fetaler Fehlbildungen unterschieden, die in den Diskussionen als atavistische Strukturen gewertet werden.
A. Häufig in der Mittellinie bzw. gering rechts oder links der Mittellinie des Rückens gelegene wurmartige Anhängsel in der Region über dem Kreuz- und Steißbein (über 95% der beschriebenen Fälle), seltener über der Lendenwirbelsäule, mit einem Durchmesser von 0,5-3,0 cm und einer Längenvarianz von 2-20 cm (max. 35 cm – aus unsicherer Quelle). Die strukturbildenden Gewebeanteile sind Fett, Bindegewebe und Gefäße. Sehr selten finden sich Muskelzellen, niemals Knochen. In der Tiefe besteht in der Regel eine Beziehung von Gewebezügen zur Wirbelsäule bzw. der Hirnhaut des Rückenmarks. Häufig zeigt die umgebende Haut eine deutliche Behaarung. Die Auswüchse zeigen unbehandelt eine Wachstumstendenz von 1-5 mm pro Woche in den ersten Lebensmonaten (Islam 2002, Gaskill & Marlin 1989; vgl. Abb. 1).
 |
 |
| Abb. 2: „True Tails“ Variante B: isolierte Anlage vergrößerter Steißbeinwirbelkörper (rechts) oder kegelförmige Wucherungen mit Kontakt zur Steißbeinspitze (links). (Nach Fara 1977 und Bar-Maor et al. 1980). |
|
B. Isolierte Anlage von drei bis fünf der sonst verschmolzenen Steißbeinwirbel, die zusätzlich eine deutliche Vergrößerung (Hypertrophie) und dadurch bedingte überdurchschnittliche Längenzunahme des Steißbeins mit entsprechender kegelförmiger Vorwölbung der Haut von max. 3-5 cm über der Steißbeinspitze aufweisen können. Dazu zählen auch Knorpelgewebe enthaltende oder bindegewebige Anhängsel bzw. kegelförmige Wucherungen, die sich direkt als untere Fortsetzung der Steißbeinspitze darstellen (Fara 1977, Bar-Maor et al. 1980; vgl. Abb. 2).
C. Gegenüber den unter A und B genannten krankhaften atypischen Bildungen wird häufig ein regelhaft auftretender ontogenetischer Merkmalskomplex als echte Rekapitulation* von Schwänzen menschlicher Vorfahren gewertet (z.B. bei Kunimoto 1918, Haeckel 1877, Baruchin et al. 1983).
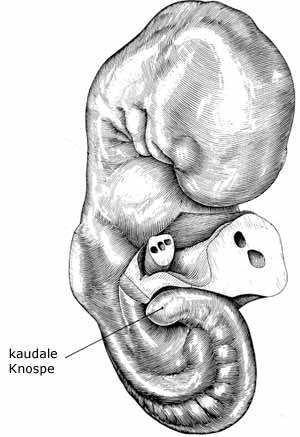
Dabei handelt es sich um die von der 4. bis 8. Entwicklungswoche beim menschlichen Embryo erscheinende kaudale* Knospe oder kaudale Eminenz („caudal bud“), die in diesen vier Entwicklungswochen einen rasanten Strukturwandel erfährt und die unterste Rumpfgrenze darstellt. Das heißt, sie überragt nur in dieser Zeit die zukünftige Öffnung des After. In der 4. Woche wird diese fast ausschließlich durch die Chorda neuralis (ein relativ fester Stab aus neuroektodermalem Gewebe) gebildet. In ihrem Umfeld bilden sich dann von (proximal nach distal) in der 5. Entwicklungswoche segmental* angeordnete Somiten und daraus die knorpligen Anlagen des Kreuz- und Steißbeins (= Wirbelkörperblasteme, vgl. Abb. 3 und 4; O‘Rahilly & Müller 1999).
 |
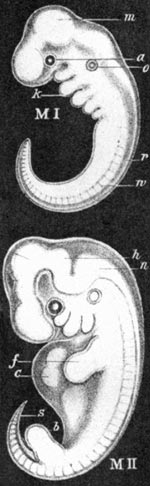 |
Abb. 3 (links): „True Tails“ Variante C: Embryo im Alter von 4 1/2 Wochen, die kaudale Knospe bildet das untere Körperende. (Nach Blechschmidt)
Abb. 4 (rechts): „True Tails“ Variante c: menschliche Embryonen nach Haeckel 1874, der das untere Körperende deutlich verlängert sowie durchgängig segmentiert darstellt (bewußte Falschdarstellung) und als „Schwanz“ bezeichnet.
|
|
|
|
Geschichtlicher Rückblick
 |
| Abb. 5: Griechischer Gott Satyr mit haarigem Büschelschwanz. (Aus Williams & Leavitt 2004) |
|
Die Vorstellung, daß auch Menschen Tierschwanzrudimente aufweisen, ist schon seit der Antike bekannt. Den griechischen Gott Pan und seine römischen Ableger Satyr und Silens stellte man als menschliche Gestalten mit einem in Höhe der mittleren Lendenwirbelsäule entspringenden büschelartigen (Haar-) Schwanz dar (Gaskill & Marlin 1988, vgl. Abb. 5). Bis heute gelten im indopersischen Kulturgebiet Kinder mit den unter A genannten Fehlbildungen als Reinkarnationen eines hinduistischen Gottes. Wissenschaftlich diskutiert wurde das Thema eines „human tail“ zuerst im Zusammenhang mit ontogenetischen Befunden. Nach Kunitomo (1917) basierte unter anderen Darwins Darstellung der Abstammung des Menschen von einer schwanztragenden äffischen Rasse auf Illustrationen des menschlichen Embryos von Ecker (1851-1859), der den unteren (= kaudalen) Anteil als „schwanzförmiges Körperende“ bezeichnete.
Darwin selbst ging bekanntlich von einer engen Wechselbeziehung zwischen der Individualentwicklung der Organismen und ihrer Stammesgeschichte aus. Seine Vorstellungen basierten auf Angaben von v.Baer, die Darwin in eine phylogenetische Lesart übertrug: „Die Abstammung ist aus meiner Sicht das geheime verbindende Band, das die Naturalisten im Begriff des natürlichen Systems gesucht haben. Aus diesem Blickwinkel können wir verstehen, daß für die meisten Naturalisten bei der Klassifikation die Struktur des Embryos wichtiger ist als die des ausgewachsenen Individuums. Denn der Embryo repräsentiert das Lebewesen in seinem am geringsten modifizierten Stadium und offenbart insofern das Bild seines Vorfahren“ (Darwin 1859, S. 449, zit. nach Richards 1992). Ins Extrem gesteigert finden wir diesen Zusammenhang bei Ernst Haeckel. Seine Embryonenbilder zeigen menschliche Embryonen mit deutlich – aber künstlich! – verlängertem kaudalem Pol, den er Schwanz nannte (Anthropogenie 1874, Abb. 4).
 |
| Tab. 1 (oben): Klassifikation kaudaler Anhänge nach Bartels (1884) in fünf Typen auf der Basis von 126 übermittelten Einzelfällen.
Tab. 2 (unten): Klassifikation kaudaler Anhänge nach Virchow (1880).
|
|
Eine der ersten wissenschaftlichen Beschreibungen einer schwanzartigen Fehlbildung bei einem neugeborenen Menschen präsentierte 1881 Miller. Er wertete sie wie viele nachfolgende Autoren als Beleg für die Richtigkeit der Rekapitulationstheorie, obwohl ein ontogenetischer Zusammenhang zwischen den bei Haeckel dargestellten embryonalen Strukturen mit den beschriebenen wurmartigen Anhängen bis dahin niemals belegt werden konnte. So klassifizierte z.B. 1884 Bartels fünf Typen kaudaler Anhänge (vgl. Tab. 1) anhand von 126 übermittelten Einzelfällen unter der Vorgabe, daß die Rückbildung der embryonalen Schwanzentwicklung des Menschen auf verschiedenen Stufen zum Stehen kommen könne. Virchow (1880) lehnte die Klassifikation Bartels‘ ab und schlug stattdessen vor, nur dann von einem „Schwanz“ beim Menschen zu reden, wenn tatsächlich den Tierschwänzen entsprechende Bildungen nachweisbar werden (Typ 1 nach Bartels). Er schlug deshalb die in Tab. 2 wiedergegebene Einteilung vor.
1901 erfolgt durch Harrison ein weiterer Klassifikationsversuch, der von der Annahme ausging, daß die Anhänge aus dem mesodermalen* Anteil (dem mittleren Keimblatt des Embryos) der embryonalen Schwanzanlage des Menschen hervorgehen. In den folgenden Jahrzehnten gab es zahlreiche Versuche, Kriterien zu definieren, um entsprechende Fehlbildungen den „echten Schwänzen“ oder den „Pseudotails“ zuzuordnen.
Der bereits erwähnte Kunimoto bemühte sich in einer 1918 publizierten sehr ausführlichen Arbeit um die Begründung einer realen Existenz des Schwanzes während der Embryonalentwicklung des Menschen. Bei den Entwicklungsbiologen stand jedoch aufgrund ihrer deutlich stärker ausgeprägten Distanz gegenüber dem Rekapitulationsdenken Haeckels in der Folge die Frage im Mittelpunkt, ob die Schwanzbildung generell bei allen Wirbeltieren nach einem gleichartigen oder nach unterschiedlichen Prinzipien verläuft. Während Holmdahl (1925) diese als einen der Gliedmaßenentwicklung analogen Vorgang betrachtete (im Rahmen der sogenannten „sekundären Körperentwicklung“), machte Vogt (1926) die gleichen Prozesse verantwortlich, die der Kopf- und Rumpfentwicklung (die sogenannte „primäre Körperentwicklung“) zugrunde liegen. Die Diskussionen sind bis heute noch im Gange, ob die embryonale Knospe durch die primäre oder sekundäre Neurulation* entsteht. Aus der Beantwortung dieser Frage ergeben sich unterschiedliche Konsequenzen für die Stammbaumrekonstruktion der Wirbeltiere (vgl. dazu Handrigan 2003).
|
Atavismen: „Rückschläge“. Strukturen, die als das Wiedererscheinen eines in der Stammesgeschichte verlorengegangenen Einzelmerkmals betrachtet werden. Ursächlich vermutet werden meist Mutationen oder genetische Rekombinationen, die verborgene Entwicklungsprogramme reaktivieren. histologisch: den Aufbau eines Gewebes betreffend. homolog: bauplangleich; in diesem Beitrag auch im Sinne von „abstammungsverwandt“. Hox-Gen-Cluster: Besondere Gruppe von Genen, die Eiweiße für die Steuerung von Transkriptionsprozessen (Übersetzung der genetischen Information) codieren. Die Reihenfolge ihrer Anordnung auf den Chromosomen entspricht z.T. der Reihenfolge ihrer Aktivierung in den jeweiligen Körperachsen. Sie besitzen eine große Relevanz für aktuelle phylogenetische Diskussionen, weil nahezu identische Grundsequenzen mit entsprechender funktioneller Bedeutung bei allen Organismen auftreten. kaudal: anatomisch-topographische Richtungsbeschreibung: fußwärts oder schwanzwärts. Mesoderm: aus dem mittleren Keimblatt der embryonalen Keimscheibe hervorgehendes Gewebe. Neurulation: ontogenetische Bildung des Neuralrohres, aus dem später die Gewebe und Strukturen des zentralen Nervensystems mit Gehirn und Rückenmark sowie die umgebenden Hirnhäute hervorgehen. Ontogenese: Gesamtheit der vorgeburtlichen Individualentwicklung, wird beim Menschen unterteilt in die Embryogenese (bis zum Ende der 8. Entwicklungswoche) und die Fetogenese (9. Entwicklungswoche bis zur Geburt). Primäre Neurulation: Das Neuralrohr entsteht durch einen Faltungsprozeß des äußeren Keimblattes (Ektoderm); typisch für Kopf und Rumpfregion bei Säugetieren. Pseudotail: Gruppe vielfältiger Mißbildungen im Bereich des unteren Rückens, die klinisch in Kombination mit ausgeprägten bösartigen Tumorbildungen oder schweren Entwicklungsstörungen der Wirbelsäule und/oder des Zentralen Nervensystems auftreten. Rekapitulation: ontogenetisch auftretende Strukturen eines Individuums, die ausgebildeten oder embryonalen Merkmalen stammesgeschichtlicher Vorfahren als homolog zugeordnet werden. Die Vorstellung basiert auf der Idee Haeckels, daß die Individualentwicklung eines Individuums die Wiederholung seiner stammesgeschichtlichen Herausbildung repräsentiert (Rekapitulationstheorie). segmental: regelhafte Aufeinanderfolge ähnlicher (jedoch nicht immer identischer) Bausteine zu einem Strukturgefüge (z.B. Somiten beim Embryo, Wirbelkörper beim Erwachsenen). Sekundäre Neurulation: Das Neuralrohr entsteht durch einen Aushöhlungsprozeß eines aus neuroektodermalem Gewebe bestehenden Stabes, typisch für die Anteile des Rückenmarks unterhalb der Lendenwirbelsäule. Teratologie: Mißbildungslehre. Teratologische Merkmale sind Fehlbildungen von Strukturen oder Organen als Folge von Entwicklungsstörungen unterschiedlichen Schweregrades.
|
|
|
Anatomie und Funktion von echten Schwänzen
Um zu bewerten, ob die angesprochenen Fehlbildungen Atavismen oder die ontogenetisch auftretende kaudale Knospe Rekapitulationen repräsentieren, sind zunächst einige Anmerkungen zur Anatomie und der Funktion echter Schwänze notwendig. Danach werden die Prinzipien der ontogenetischen Entwicklung des Körperendes bei den Wirbeltieren und der des Menschen vorgestellt. Die bekannten Ursachen der Fehlbildungen sind Inhalt des darauffolgenden Abschnitts. Auf diesen Grundlagen erfolgt dann die Analyse und Beurteilung von Homologiebetrachtungen*.
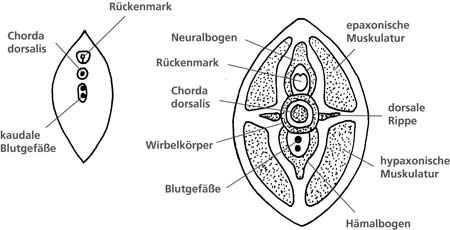 |
| Abb. 6: Einfacher und komplexer Grundbauplan des Schwanzes der Wirbeltiere. (Nach Romer & Parsons 1991) |
|
Der Schwanz ist ein charakteristisches Merkmal nahezu aller Organismen, die zu den Wirbeltieren (Vertebraten) zählen. Damit unterscheiden sie sich u.a. von den übrigen Unterstämmen der Chordata (Tiere mit Anlage einer Chorda dorsalis), zu denen z.B. die Tunicaten (Manteltiere) und das Lanzettfischchen (Amphioxus) zählen. Bei ausgewachsenen Wirbeltieren läßt sich ein allgemeiner Bauplan des Schwanzes wie folgt beschreiben (nach Romer & Parsons [1991], vgl. dazu Abb. 6). Im Querschnitt zeigt dieser typischerweise ein aufrecht stehendes Oval, zentral gelegen ist die Wirbelsäule – speziell die Elemente des Steißbeins (Os coccygis) –, darüber angeordnet ist das Neuralrohr, in dem Nerven, Anteile des Rückenmarks oder nur die Rückenmarksflüssigkeit zu finden sind. Jegliche Andeutung einer Art Leibeshöhle mit den von ihr umschlossenen Eingeweiden fehlt. Weiterhin zur Grundstruktur gehören segmental angeordnete Muskelgruppen und unterhalb der Wirbelkörper verlaufende Blutgefäße. Bei den Wirbeltieren besteht eine große Variabilität bezüglich der Anzahl der Wirbelkörper (30-200), bei den meisten Säugetierordnungen hingegen ist bis auf die Schwanzregion (0-50 coccygeale Wirbelkörper) die Anzahl der übrigen Wirbelkörper im Bereich der Hals-, Brust-, Lendenwirbelsäule und des Kreuzbeins relativ konstant.
Eine Ausnahme von diesem Grundbauplan zeigt die Makakenaffenart M. sylvanus (Berberaffe), dessen fleischiger Schwanz keine Wirbelkörper enthält.
Die Funktionalität des Schwanzes ist sehr vielfältig. Er dient unter anderem der Fortbewegung, der Balance, der Stabilisierung und Steuerung von Bewegungen, als Greiforgan, Schmuck, Verteidigungswaffe oder Schutzorgan.
|
|
Grundzüge der ontogenetischen Entwicklung der Steißbeinregion des Menschen
Grundsätzlich entsteht der Schwanz bei Wirbeltieren aus der embryonalen Anlage der kaudalen Knospe (Schwanzknospe), die sich ihrerseits im Zusammenhang mit der Chorda neuralis und einer besonderen Form der Neurulation* bildet. Die anschließende ontogenetische Bildung des Steißbeins mit den Schwanzwirbeln zeigt deutliche Besonderheiten im Vergleich zur Entstehung der übrigen Wirbelkörper. Dies soll am Beispiel des Menschen kurz erläutert werden.
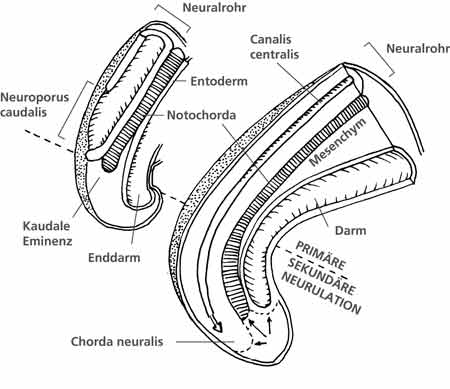 |
| Abb. 7: Entwicklung der kaudalen Knospe beim Menschen. (Nach O'Rahilly & Müller 1999) |
|
In der 4. Entwicklungswoche wird bei dem ca. 2,5-4,5 mm großen menschlichen Embryo die Schwanzknospe sichtbar. Ein stabartiger Auswuchs (Chorda neuralis) des im embryonalen Rumpf bereits angelegten Neuralrohres beginnt ab Höhe des Neuroporus caudalis in Richtung der zunächst allein aus mesodermalen Zellen bestehenden kaudalen Knospe auszuwachsen. Dabei überwächst diese Chorda neuralis das untere Ende der Chorda dorsalis (ein knorpliger Stab, der u.a. die Wirbelkörperbildung oberhalb des Kreuzbeins induziert) sowie das Ende des Enddarms (Anus). Die kaudale Knospe stellt ab der 4. bis zur der 8. Entwicklungswoche (der Embryo ist jetzt 27-31 mm groß) das untere Ende des embryonalen Körpers dar. Das Neuralrohr, aus dem später u.a. das Gehirn, das Rückenmark und die Nerven hervorgehen, bildet sich im Rumpf und im Kopf über den Weg der primären Neurulation. Dabei falten sich die Blätter des embryonalen Nervengewebes zu einem Rohr zusammen. Anders als in den übrigen kopfwärts gelegenen Abschnitten entstehen innerhalb der Chorda neuralis der Schwanzknospe durch innere Aushöhlung (= sekundäre Neurulation*) die unteren Anteile des Neuralrohres. Dieses gewinnt nach oben (proximal) Anschluß an den durch die primäre Neurulation gebildeten oberen Anteil des Neuralrohres. In dieser Phase wird der untere Anteil der Chorda neuralis zum Ende des gesamten Neuralrohrs und macht den Hauptbestandteil der Schwanzknospe aus.
Bei einigen Vertretern der Wirbeltiere vollzieht sich die Bildung des Neuralrohres auch in der Schwanzknospe nach dem Prinzip der primären Neurulation (so z.B. bei den Schleimfischen, den Knorpelfischen, einigen Lungenfischen). Die sekundäre Neurulation gilt nach Handrigan (2003) als phylogenetisch abgeleitete Variante. Diese Ansicht ist jedoch aufgrund der noch lückenhaften Datenlage und Problemen bei der Stammbaumrekonstruktion (z.B. vielfach anzunehmende Konvergenzen) umstritten.
Die Chorda neuralis induziert in ihrer Umgebung die Bildung von Bindegewebe und später die Anlage der von acht Steißbeinsomiten (= vom Mesenchym gebildete gegliederte Strukturen neben dem Neuralrohr), aus welchen u.a. drei bis fünf verschmolzene Steißbeinwirbel und Anteile der Beckenbodenmuskulatur hervorgehen. Diese Somiten reichen anfänglich nicht bis zum Körperende. Erst am Ende der 7. Woche wird das Neuralrohr vollständig von den Anlagen der Steißbeinsomiten umgeben, die in Relation zum Neuralrohr nun ein deutlich schnelleres Wachstum in allen drei Raumebenen aufweisen und es später deutlich überwachsen. Beim Menschen und den Menschenaffen ragen die von den Steißbeinsomiten gebildeten Strukturen nie über das untere Körperende hinaus. Bei den schwanztragenden Wirbeltieren entwickelt sich aus den Steißbeinsomiten der Schwanz mit der oben beschriebenen typischen Anatomie.
O’Rahilly & Müller (1999, 348) betonen mit Nachdruck, daß keine Ähnlichkeiten zwischen der embryonal angelegten kaudalen Knospe und einem ausgebildeten Wirbeltierschwanz bestehen, weder auf morphologischer noch funktioneller Ebene (vgl. dazu Abb. 7).
In den letzten Jahren wurde die Bedeutung der Hox-Gen-Cluster* für die Differenzierung der kaudalen Knospe und der daraus hervorgehenden Strukturen weiter erhellt. Analog der Situation bei der Gliedmaßenentwicklung zeigen die Zellen der kaudalen Eminenz anfangs einen homogenen histologischen* Aufbau, jedoch entsprechend ihrer Position in Bezug auf die Raumachsen ein genetisch differenziertes Expressionsmuster. Den Hox-Gen-Clustern 9-13 kommt eine führende Rolle bei der Steuerung der typischen Herausbildung der spezifischen Wirbelkörper entlang der Körperachse (z.B. Brustwirbelkörper mit Rippen, Lendenwirbelkörper ohne Rippen, Sakralwirbel mit verschmolzenen Querfortsätzen, Steißbeinwirbel verschmolzen ohne Rippenansatz) zu. Dabei konnte gezeigt werden, daß das jeweils distal aktiv werdende Hox-Gen-Cluster die Funktion der unmittelbar proximal gelegenen Gruppe hemmt. So verhindert z.B. Hox-10 bei Mäusen die Ausbildung von Rippen im Bereich der Lendenwirbelsäule bis zum Steißbein, Hox-11 dagegen hemmt die Wirkung von Hox -10 im Bereich der Sakralwirbel, weshalb dort die verschmolzenen Querfortsätze das typische Bild des Kreuzbeins bilden (Wellik & Capecchi 2003). Interessant ist die auch die Tatsache, daß bei Mäusen Mutationen mit einem Funktionsverlust des Hox-Gen-Clusters 13 (sog. „loss-off-function Mutation“) zu einer deutlichen Vergrößerung und numerischen Zunahme der Schwanzwirbelkörper führen, deren Verschmelzung jedoch weitestgehend ausbleibt (Economides et al. 2003, vgl. Abb. 8 und 9).
|
|
Ursachen krankhafter Entwicklungen in der Steißbeinregion des Menschen
Während die skizzierte Bildung und Umwandlung der kaudalen Knospe den normalen ontogenetischen Entwicklungsgang darstellt, sind die beim Menschen unter „A“ und „B“ bekannten embryo-fetalen Fehlbildungen Resultate krankhafter (teratologischer) Entwicklungsprozesse. Im folgenden sollen sie aus dem Blickwinkel verfügbarer pathologischer Zusammenhänge und bekannter Entwicklungsstörungen charakterisiert werden.
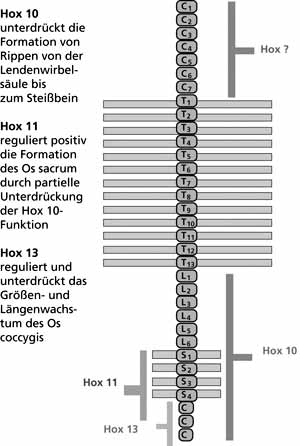 |
| Abb. 8: Bekannte Einflüsse einzelner Hox-Gen-Cluster* auf die Differenzierung der unteren Wirbelsäule bei Mäusen. (Nach Economides et al. 2003 und Wellik & Capecchi 2003) |
|
A. Die von der Mittellinie bzw. gering rechts oder links der Mittellinie ausgehenden wurmartigen Anhängsel (Abb. 1) zeigen bei exakter chirurgischer Freilegung häufig eine Verbindung zum primär gebildeten Neuralkanal, jedoch nie zum sekundär gebildeten (Gaskill & Marlin 1989). In keinem Fall wurden Anlagen von Wirbelkörpern oder das Vorhandensein von Resten einer Chorda neuralis in diesen Fehlbildungen beschrieben. Histologisch handelt es sich um Gebilde aus reifem Fettgewebe, wenigen ungeordnet strukturierten Anteilen von Muskelzellen und Gefäßstrukturen sowie anderer Bindegewebskomponenten. Entgegen anfänglicher Annahmen treten in 20-30% der als „true tails“ beschriebenen Fälle noch andere embryo-fetale Mißbildungen auf (z.B. Spina bifida – ein fehlender Verschluß der Wirbelbögen –, Tumoren wie Lipome und Hämangiome, Meningocelen – flüssigkeitsgefüllte Ausstülpungen der Rückenmarkshäute –, fehlende Anlage von Kreuzbeinwirbeln, Spaltmißbildungen des Gaumens usw.).
Für diese Fehlbildungen ursächlich verantwortlich gemacht werden können Störungen des Verschlusses des Neuralrohres, des Neuroporus inferior und/oder des Prozesses der sekundären Neurulation. Speziell das Wechselspiel von Zellneubildung und Zellrückbildung (Apoptose) in diesen sensiblen Regionen entkoppelt sich von der normalen Regulation. Gaskill & Marlin (1989) nennen diese Anhängsel deshalb „neuroektodermale Appendices“, deren Deutung als Atavismus auch aus anderem Grund abzulehnen ist. Dazu verweisen sie zusätzlich auf analoge morphologische Befunde, die an anderen Körperregionen beim Menschen auftreten können (z.B. in der Kopf-Halsregion, Oberschenkelinnenseite, Übergang von Halswirbelsäule zur Brustwirbelsäule).
B. Die zahlenmäßige Mehranlage voneinander isolierter Steißbeinwirbel, die Hypertrophie (= vermehrtes Wachstum) einzelner oder eines Steißbeinwirbels mit überdurchschnittlicher Ausdehnung in der Längsrichtung des Körpers und dadurch bedingter Vorwölbung der Haut oder die kegelförmige Wucherung an der Steißbeinspitze sind äußerst seltene Befunde im Vergleich zur Variante „A“. Nach experimentellen Ergebnissen an der Maus entsprechen diese Fehlbildungen dem morphologischen Bild bei Funktionsverlusten der Hox 10- und Hox 11-Cluster sowie der loss-off-function Mutation im Hox-b13-Cluster. In diesen Fällen trat bei den Versuchstieren ein überschießendes Wachstum der Strukturen auf, die sich aus der Chorda neuralis ableiten oder durch sie induziert werden. Die z. T. dabei beobachtete numerische Mehranlage von max. 1-2 Steißbeinwirbelkörpern beruht auf der fehlenden Verschmelzung der primär isoliert angelegten Wirbelkörperanlagen des Steißbeins und ihrer eigenständigen Differenzierung zu Wirbelkörpern. Eine weitere Ursache sind ubiquitär auftretende embryonale Tumorbildungen (Teratome), die u.a. die kegelförmigen Wucherungen an der Steißbeinspitze wie bei Fara (1977) beschrieben hervorrufen können.
|
|
Ist eine Homologisierung von Fehlbildungen und ontogenetischen Merkmalen mit echten Schwänzen gerechtfertigt?
Virchow hatte bereits 1880 gefordert, nur die Formen wurmartiger Anhängsel (Appendices) als Schwanzhomologon zu bezeichnen, welche eindeutig das entsprechende Merkmalsmuster eines Wirbeltierschwanzes aufweisen.
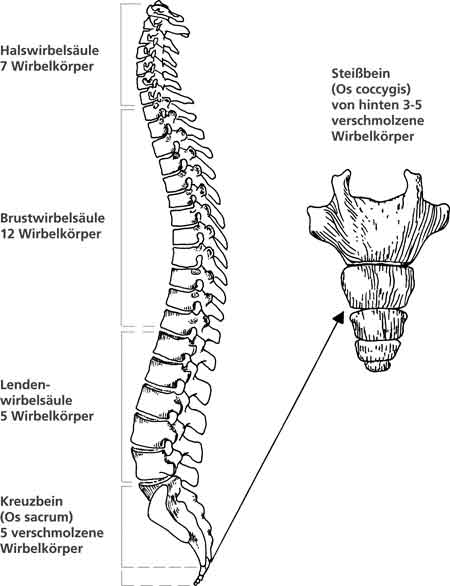 |
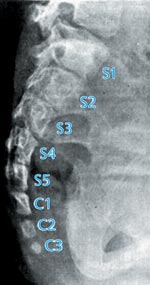
| Abb. 9: Schematische Anatomie der Wirbelsäule und des Steißbeins. (Nach Fenis 1988) |
|
Die unter „A“ dargestellten Fehlbildungen erfüllen in keinem der bisher beschriebenen Fälle die notwendigen Kriterien, um als homolog den echten Wirbeltierschwänzen gegenübergestellt zu werden. Insbesondere leiten sie sich nicht als ein unterer (distaler) Fortsatz aus der embryonal angelegten kaudalen Eminenz ab, wie es die normale Entwicklung der Schwänze bei den entsprechenden Wirbeltieren zeigt. Die Fehlbildungen entstehen in Folge o.g. Störungen im Gebiet entlang der Verschlußzone des früheren Neuralrohres, insbesondere im Bereich des Neuroporus inferior, der topographischen Grenze am embryonalen Übergang vom Rücken zum Becken. Auch entspricht ihr morphologisch-histologisches Erscheinungsbild in keinem Fall der Situation eines echten Wirbeltierschwanzes. Auch ihr nachgewiesenes Auftreten an anderen schwanzuntypischen Körperstellen spricht gegen die Vorstellung, darin atavistische „true tails“ beim Menschen zu sehen. Der Hinweis auf eine eventuelle Ähnlichkeit mit dem wirbelkörperlosen Schwanz des Makakenäffchens M. sylvanus (wie bei Theobald 2004) ist aufgrund der o.g. Zusammenhänge ebenfalls nicht haltbar, da dieser die unmittelbare Fortsetzung der Steißbeins darstellt.
Sachlich korrekt ist die Homologisierung des Steißbeins des Menschen mit den Steißbeinen schwanztragender Wirbeltiere. Doch diese Ähnlichkeit ist nicht mehr, aber auch nicht weniger bedeutsam wie die zwischen den Extremitäten oder den Schädelknochen bei Wirbeltieren.
Der Unterschied liegt bei gleichem Grundbauplan in der Anzahl und der Modifikation der gebildeten Steißbeinwirbelkörper, ihrer damit verbundenen Gesamtlänge, die über den After hinausreicht, und ihrer Funktionalität. Diese Unterschiede bleiben aber auch bei den unter „B“ beschriebenen pathologischen Veränderungen unverändert bestehen. Ein Zusammenhang zwischen zuviel angelegten Steißbeinsomiten (mehr als acht) und der Ausbildung dieses Fehlbildungstyps konnte nicht belegt werden. Die voneinander isolierten Steißbeinwirbelkörper ragen nicht als schwanzähnliche Anhängsel über den Damm hinaus, sondern wölben diesen bei entsprechender Hypertrophie nur deutlich vor. Die kegelförmige Wucherung ausgehend von der Steißbeinspitze ist ein embryonaler Tumor (Teratom), der auch in vielen anderen Körperregionen auftreten kann. Hier verbietet sich von allein jeglicher Versuch einer Homologisierung.
Bei allen Diskussionen wird eine wesentliche Frage ausgeblendet: Worin unterscheidet sich ein atavistisches Merkmal gegenüber den teratologischen Folgen sonstiger Mutationen mit entsprechend veränderter Merkmalsausprägung?
Wenn ein Atavismus angenommen wird, dann sollte der mögliche Vorfahr, dessen Merkmal beim Menschen konserviert blieb, benannt werden können. Hier ist die Sachlage auch nicht einfacher. Gängige evolutionstheoretische Vorstellungen sehen in den afrikanischen miozänen schwanzlosen Menschenaffen (lebten im Zeitraum von vor 22 -5 Millionen Jahren) wie Proconsul die Vorfahren des Menschen (Begun 2004). Von welcher Form der Altweltaffen diese wiederum abstammen und über welche Entwicklungswege die Evolution zum Menschen verlief, ist nach wie vor offen. Mit welcher Sicherheit kann dann überhaupt von einem schwanztragenden Vorfahren des Menschen gesprochen werden?
Die embryonale Anlage des unteren Körperendes bei den Wirbeltieren, die kaudale Knospe, ist bei allen Wirbeltieren unter dem Blickwinkel einer entsprechenden ontogenetischen Entwicklungsphase homologisierbar. Diese embryonalen Strukturen stellen aber aus morphologischer und funktioneller Perspektive (O’Rahilly & Müller 1999) keine homologe Entsprechung zu den adulten (ausgewachsenen) Wirbeltierschwänzen dar (vgl. die Interpretation des im ersten Abschnitt genannten Phänomens „C“). Die kaudale Knospe wird hauptsächlich durch die unsegmentierte Chorda neuralis und das sekundär gebildete Neuralrohr gestaltmorphologisch charakterisiert und nicht durch segmentale angeordnete Wirbel und willkürlich bewegbare Muskelstrukturen. Sie bildet die Matrix, aus der sich neben den Steißbeinwirbeln beim Menschen und den Schwanzwirbeln bei den entsprechenden Wirbeltieren auch Anteile des knöchernen Beckens und seiner Weichteilstrukturen ableiten. Letztere würden bei einer Homologisierung der Schwanzknospe lediglich mit den Wirbeltierschwänzen völlig ausgeklammert.
Die zur Diskussion gestellten Phänomene (A: wurmartige Anhängsel im Bereich des Steißbeins, B: isolierte oder tumoröse Vergrößerung einzelner Steißbeinwirbel, C: embroynal vorübergehend auftretende Schwanzknospe) beim Menschen, die als Atavismen oder Rekapitulation von Schwänzen interpretiert werden, zeigen deutlich die Schwäche und methodologische Unschärfe des phylogenetischen Homologieverständnisses. Die embryonale Struktur rekapituliert nach dieser Lesart die noch vorhandene Schwanzanlage unserer vermeintlichen Vorfahren. Auch die beschriebenen Fehlbildungen gelten als Atavismen ehemaliger Schwänze. An welcher Stelle wird nun aber eigentlich richtig rekapituliert und wo haben wir es hingegen mit einer nicht-rekapitulierten Ähnlichkeit zu tun? Diese Entscheidung trifft der Forscher allein auf der Grundlage subjektiver Erwägungen.
 |
| Der suggestive Terminus „Schwanz“ und ähnliche Begriffe haben für die Fehlbildungen beim Menschen keine Berechtigung. |
 |
|
Der suggestive Terminus „Schwanz“ oder „human tail“ sowie andere abschwächende Begriffe („pseudotail“) haben für die besprochenen Fehlbildungen beim Menschen keine Berechtigung. Auch die Darstellung der embryonalen kaudalen Eminenz als Anklang eines ehemals bei den Vorfahren vorhandenen Schwanzes ist nicht begründet. Neben den vorhandenen anatomisch-funktionellen Fakten und den Daten der vergleichenden Ontogenese spricht die angedeutete methodische Schwäche von Homologiebetrachtungen und die schon an anderer Stelle (Ullrich 1994) diskutierten ungeklärten kausalen Mechanismen einer Verknüpfung von Ontogenese und Phylogenese gegen eine solche Argumentation. Nicht zuletzt auch deshalb wird der Wert der vergleichenden Anatomie und Embryologie für die Erhellung von Stammbaumbeziehungen mehr und mehr von Evolutionstheoretikern kritisch hinterfragt (Richardson & Verbeek 2003).
|
In einigen Fällen von Diastematomyelie, wo das Rückenmark durch eine Fehlbildung in zwei Strängen im Bereich der Brust- und Lendenwirbelsäule angelegt ist, treten an der Haut über der Lendenwirbelsäule einzelne büschelartige Haarauswüchse auf, die eine beträchtliche Länge (über 20 cm) erreichen können. Es ist nach Williams & Leavitt (2004) als sicher anzunehmen, daß diese Fehlbildung die intuitive Vorlage für den Entwurf des griechischen Gottesbildes von Pan ca. 500 Jahre vor Christus war. Die Autoren gehen davon aus, daß offenbar das ungewöhnliche Erscheinungsbild eines Erkrankten (zusätzlich tritt als Symptom bei Diastematomyelie eine Spitzfußstellung auf, Pan wird immer mit einem Huf-Fuß dargestellt!) die Einstufung jener Menschen als eine Art göttliches Mischwesen zwischen Tier und Mensch hervorrief.
Dieses illustrative Beispiel und die dargestellten Diskussionen um die Existenz eines „human tail“ zeigen erneut den enormen Einfluß weltanschaulicher oder religiöser Grundpositionen auf die Beschreibung und Interpretation von Fakten – ein Zusammenhang, dem jeder Wissenschaftler unterworfen ist.
|
|
Literatur
- Bar-Maor JA et al. (1980)
- Human tails. J. Bone Joint Surg. 62B, 508-510.
- Baruchin AM et al. (1983)
- The human caudal appendage (human tail). Brit. J. plast. surg. 36, 120-123.
- Bartels M (1884)
- Die geschwänzten Menschen. Arch. Anthropol. 15, 45.
- Begun DR (2004)
- Das Zeitalter der Menschenaffen. Spektr. Wiss. Dossier 1-04, 6-14.
- Dao AH (1983)
- Human tails and pseudotails. Hum. Pathol. 15, 449-453.
- Dubrow TT (1988)
- Detailing the Human Tail. Ann. Plast. Surg. 20, 340-344.
- Ecker A (1851-1859)
- Icons Physiologica. Erläuterungsgeschichte. Leipzig.
- Economides KD et al. (2003)
- Hoxb13 mutations cause overgrowth of caudal spinal cord and tail vertebrae. Developmental Biology 256, 317-330.
- Fara M (1977)
- Coccygeal („Tail“) projection with cartilage content. Acta chir. plast. 19, 50-54.
- Feneis H (1988)
- Anatomisches Bildwörterbuch. Stuttgart. 6. Auflage.
- Galizia G (2001)
- True tail dorso-lumbar: malformation or ancestral remaining. Minerva Pediatr. 53, 367-370.
- Gaskill SJ & Marlin AE (1989)
- Neuroectodermal Appendages: The human tail explained. Pediatr. Neurosci. 15, 95-99.
- Haeckel E (1877)
- Anthropogenie oder Entwickelungsgeschichte des Menschen. Leipzig, 3. Auflage.
- Handrigan GR (2003)
- Concordia discors: duality in the origin of the vertebrate tail. J. Anat. 202, 255-267.
- Holmdahl DE (1925)
- Die erste Entwicklung des Körpers bei den Vögeln und Säugetieren. Gegenbaurs Morph. Jahrb. 55, 112-208.
- Islam MK (2002)
- Child with a Tail. Indian J. Pediatr. 69, 819-820.
- Kunitomo K (1918)
- The development and reduction of the tail and of the end of the spinal cord. Contribution to embryology. No. 26, 161-198.
- Miller CH (1881)
- Tailed humanity. Med. Surg. Reporter 46, 165.
- O’Rahilly R & Müller F (1999)
- Embryologie und Teratologie des Menschen. Bern.
- Richards RJ (1992)
- The meaning of Evolution. The morphological and ideological Reconstruction of Darwins Theory. Chicago, London.
- Richardson MK & Verbeek FJ (2003)
- New directions in comparative embryology and the nature of developmental characters. Anim. Biol. 53, 303-311.
- Romer AS & Parsons TS (1991)
- Vergleichende Anatomie der Wirbeltiere. 5. Auflage. Hamburg.
- Theobald D (2004)
- 29 + Evidences for Macroevolution. www.talkorigins.org/faqs/comdesc/section2.html.
- Ullrich H (1994)
- Homologie und Embryologie. Die Reichert-Gauppsche Theorie. Stud. Int. J. 1, 15-24.
- Virchow R (1880)
- Über die Schwanzbildung beim Menschen. Arch. Pathol. Anat. 79, 176.
- Vogt W (1926)
- Über Wachstum und Gestaltungsbewegungen am hinteren Körperende der Amphibien. Anat. Anz. 61, 62-75.
- Wellik DM & Capecchi MR (2003)
- Hox 10 and Hox 11 Genes are required to globally pattern the mammalian skeleton. Science 301, 363-367.
- Williams GP & Leavitt L (2004)
- How Pan got his tail. Neurology 62, 519-522.
|
|
|  |

