 |
Fleckige Flügel für Fruchtfliegen
Evolution komplexer Eigenschaften: veränderte Regulation oder echter Neuerwerb? von Niko Winkler
Studium Integrale Journal
14. Jahrgang / Heft 1 - April 2007
Seite 20 - 25
|
 |
Zusammenfassung: Verschiedene Fruchtfliegenarten haben unterschiedliche Flügelmuster. Bisher fraglich war, wie diese Muster entstehen; insbesondere dann, wenn nahe verwandte Arten nur durchsichtige, ungemusterte Flügel besitzen. Es konnte gezeigt werden, dass kleine genetische Änderungen in den Regulationsbereichen eines farbgebenden Gens dafür verantwortlich sind. Die grundsätzliche Frage nach der Entstehung der zugrunde liegenden regulativen Netzwerke wird dadurch allerdings nicht beantwortet.
| |
|
Die Regulation von Farbigkeit
Das Aussehen von Organismen wird unter anderem von ihren Genen kontrolliert. So sind beispielsweise Wellensittiche in ihrer australischen Heimat normalerweise grün. Ihre Federn enthalten sowohl einen blauen als auch einen gelben Farbstoff. Fällt Blau aus, so erscheinen die Tiere gelb, fällt Gelb aus, so erhalten wir blaue Wellensittiche. Daneben sind die dunklen Federn mit einem weiteren Farbstoff, Melanin, farblich abgesetzt. Beim Ausfall aller drei Farben erhält man reinweiße Tiere. Die Veränderung solcher Farbmuster ist in der Natur jedoch eher selten auf einen Totalausfall (Verlustmutation) zurückzuführen. Meistens sind die Farb- oder Musterabänderungen graduell und gehen auf eine räumlich veränderte Gen-Ausprägung zurück.
Man kann, in vereinfachter Form, Verlust- und Regulationsmutanten des Farbstoffs Melanin bei Wirbeltieren an deren Augen unterscheiden: Echte Albinos können überhaupt kein Melanin mehr produzieren und haben daher auch rote Augen (Verlustmutante). Bei Tieren mit schwarzen Augen, aber weißen Federn oder Fell ist lediglich die Melaninproduktion in den Federn bzw. im Fell herunterreguliert oder abgeschaltet (Regulationsmutante). Ein bekanntes, ja fast schon berühmtes Beispiel für eine Genregulation der Melaninproduktion ist der Birkenspanner. Es handelt sich hier um einen Falter, dessen Flügel normalerweise eine weiße Grundfarbe haben und mit braunen Flecken besprenkelt sind. Es sind aber auch völlig schwarze Formen sowie alle Übergänge von hell nach dunkel beschrieben worden (Eckstein 1930). Auch wenn der genetische Mechanismus in diesem Fall nicht bekannt ist, so ist doch zu vermuten, dass es sich „lediglich“ um eine jeweils veränderte „Schalterstellung“ für braune Flecken handelt, angefangen von kleinen Einsprengseln bis hin zu zusammenfließenden Flecken, die alles Weiß abdecken.
Das für die Farbausprägung verantwortliche Gen kommt also in verschiedenen Ausprägungen vor. Diese Varianten nennt man Allele und demzufolge spricht der Biologe bei einer Häufigkeitsänderung bestimmter Farbtypen von Allelfrequenzverschiebung.
|
|
Flügelflecken bei Fruchtfliegen
 |
| Abb. 1: Eine Zusammenstellung von Flügeln verschiedener Fruchtfliegen-Arten. Die Färbungen spielen beim Brauttanz der Männchen eine wichtige Rolle, die ihre Flügel den Weibchen präsentieren. (© Nicolas Gompel, Abdruck mit freundlicher Genehmigung) |
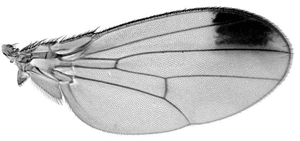 |
| Abb. 2: Flügel mit Flügelfleck, wie er z.B. bei Drosophila biarmipes vorkommt. (Ausschnitt aus Abb. 1, © Nicolas Gompel, Abdruck mit freundlicher Genehmingung) |
|
Ein beliebtes Untersuchungsobjekt für solche genetischen Fragestellungen sind die Fruchtfliegen wie Drosophila melanogaster und ihre verwandten Arten. Glücklicherweise haben Drosophila-Arten nicht nur Unterschiede in den Flügelmustern, sondern sind genetisch auch hervorragend untersucht. Die Flügel der Fliegenmännchen sind häufig auffällig gefärbt (Abb. 1). Diese Färbung spielt beim Werben um Weibchen eine Rolle, denn die Männchen umwerben die Fliegen-Weibchen vor der Begattung mit einem komplexen Brauttanz. Dabei wedeln sie unter anderem mit ihren Flügeln „vor der Nase“ der Weibchen herum, um auf sich aufmerksam zu machen. Die Flügel der Weibchen hingegen sind meist weniger prächtig oder sogar nur einfarbig. Nahe verwandte Arten haben zum Teil deutlich unterschiedliche Flügelmuster. Dies nutzten Forscher, um Licht in die Entstehung verschiedener Flügelmuster zu bringen.
Unterschiedliche Muster entstehen zuerst einmal durch unterschiedliche Genaktivitäten, die durch sogenannte cis- und trans-Elemente reguliert werden (siehe Kasten). Bei den Flügeln der Fruchtfliegen bestimmt die Menge eines Proteins mit dem Namen Yellow die Färbung. „Yellow“, weil der Ausfall des Proteins helle Fliegen entstehen lässt, die gelblich aussehen. Wenig Yellow in den Flügeln ergibt helle Stellen, viel Yellow dunkle Stellen. Die Menge an Yellow wird eben über besagte cis- und trans-Elemente gesteuert. cis-Elemente liegen dabei direkt vor dem jeweiligen Gen und steuern dieses durch die unterschiedliche Anbindung von trans-Elementen. Trans-Elemente wirken auf viele verschiedene Gene gleichzeitig. Schon lange wurde vermutet, dass gestaltliche Änderungen mit veränderten cis-Elementen einhergehen, denn Veränderungen in diesen Elementen betreffen in der Regel nur das von ihnen gesteuerte Gen. Solche Veränderungen sind daher weniger schädlich als die Veränderung von trans-Elementen, die eine Vielzahl von verschiedenen Genen steuern. Es gibt jedoch nur eine geringe Anzahl von wissenschaftlichen Arbeiten, die diese Fragen im Detail angehen. Daher war es lange Zeit unbekannt, ob die Evolution neuer gestaltlicher Eigenschaften (hier die Färbung) tatsächlich durch die Veränderung bestehender oder die Entwicklung neuer cis-Elemente geschieht. Unverstanden war weiterhin, wieviel Änderungen in einem regulatorischen Element notwendig sind, um ein neues Muster zu erzeugen.
Um diese Fragen anzugehen, untersuchten Gompel und Kollegen (Gompel et al. 2005) die nahe verwandten Fruchtfliegen D. melanogaster, D. biarmipes und D. pseudoobscura. Dabei besitzt nur D. biarmipes einen Flügelfleck auf der Spitze (Abb. 2), die beiden anderen Arten haben einfarbig durchsichtige Flügel (Abb. 1, 6. Zeile, Mitte). Die Forscher konnten bestätigen, dass in D. biarmipes (mit Flügelflecken) dort, wo später der Flügelfleck sein wird, während der Flügelentwicklung viel Yellow zu finden war. In den Arten mit durchsichtigen Flügeln war nur eine gleichmäßig verteilte, geringe Menge an Yellow zu finden. Um nun herauszufinden, ob die unterschiedlichen Flügelmuster der beiden Arten auf Veränderungen der cis- oder trans-Elemente zurückzuführen sind, fügten die Forscher die regulatorische Region des yellow-Gens von D. biarmipes (mit Flecken) in D. melanogaster (Fliege ohne Flügelflecken) ein. In der Tat entsprach nun das Muster der Yellow-Verteilung in etwa dem, wie es in D. biarmipes auftritt: eine Hintergrundaktivität verteilt über den ganzen Flügel mit einer stärkeren Konzentration an der Flügelspitze. Das bedeutet, dass die Veränderung der Färbung (= Bildung von Flügelflecken) vorwiegend auf die Veränderung von cis-Elementen zurückzuführen ist.
Mit weiteren ähnlichen Versuchen gelang den Forschern, das cis-Element, welches für den Flügelfleck notwendig ist, weiter einzugrenzen. Ein 675 Basenpaare langes Fragment, ca. 1000 Basenpaare vor dem Protein-kodierenden Bereich des yellow-Gens, ist im Prinzip ausreichend, um einen Farbfleck hervorzurufen. Dieses cis-Element wurde spot genannt. Durch weitere Serien von Versuchen konnte herausgefunden werden, dass mindestens eine Bindestelle für ein aktivierendes und eine für ein unterdrückendes trans-Element zu den bisher vorhandenen cis-Elementen hinzugekommen sind. Als unterdrückendes Element konnte nachfolgend der trans-Faktor Engrailed ausfindig gemacht werden. Dieser Faktor ist wichtig für die Festlegung von vorne und hinten. Er wird im Flügel nur in der unteren Flügelhälfte ausgebildet und teilt den Flügel so in eine vordere und hintere Hälfte. Engrailed unterdrückt yellow in der unteren Flügelhälfte, so dass der Flügelfleck eben nur im vorderen Bereich des Flügels auftritt. Der positive Faktor für die stärkere Ausbildung von yellow in der Flügelspitze wurde in der Publikation nicht näher beschrieben. Abb. 3 gibt einen vereinfachten grafischen Überblick über die Situation.
|
|
|
Evolutionäre Flickschusterei?
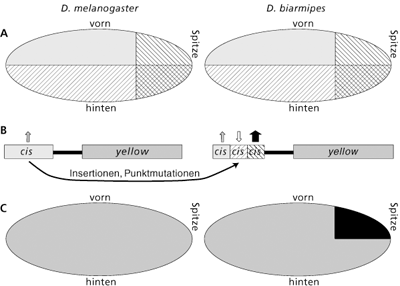 |
| Abb. 3: Entstehung des veränderten cis-Elements „spot“ bei D. biarmipes aus einem spot-Element von D. melanogaster. A Das verborgene Flügelmuster zweier trans-Elemente ist in beiden Arten gleich. B Das cis-Element vor dem Gen yellow sorgt für eine gleichmäßig schwache Ausprägung des Proteins Yellow im Flügel. Durch Mutationen verändert sich das cis-Element, so dass es von zwei weiteren trans-Elementen erkannt wird. C Ein gleichmäßig leicht grau gefärbter Flügel ergibt sich durch eine entsprechend niedrige Konzentration an Yellow in D. melanogaster. In D. biarmipes wird Yellow durch das trans-Element in der Flügelspitze stark aktiviert, in der unteren Flügelhälfte durch ein anderes trans-Element aber stark unterdrückt. In der Folge entsteht ein lokaler Farbfleck. |
|
Die beobachtete Veränderung im sichtbaren Flügelmuster (d.h. zusätzliche Flügelflecken) ist – den Autoren der Studie zufolge – durch eine so-genannte „Kooption“ zustande gekommen: Im Flügel befindet sich bereits während der Entwicklung des Insekts ein nicht sichtbares Muster aus mindestens zwei trans-Faktoren, von denen einer als Engrailed identifiziert wurde (s. o.). Dieses verborgene Muster der beiden trans-Faktoren wird von yellow „übernommen“. Das heißt, das Gen yellow hat zwei zusätzliche Bindestellen (durch Mutationen) erhalten, die nun von den beiden bereits vorhandenen trans-Faktoren erkannt werden. Da das entstehende Protein Yellow für die Entwicklung von Melanin (= dunkle Farbe) zuständig ist, wird das bisher verborgene Muster der beiden trans-Faktoren (Vormuster, „pre-pattern“) auf den Flügeln sichtbar.
Interessanterweise wird an dieser Stelle eine weitreichende Spekulation vorgestellt: So sollen Proteinbausteine, die die Linsen der Wirbeltiere aufbauen (Linsencrystalline) ebenfalls durch einen solchen „Kooptions“-Prozess als Linsenbaumaterial herangezogen worden sein. Denn die Eiweiße, die unsere Augenlinse aufbauen, findet man an anderer Stelle im normalen Zellstoffwechsel als normale Enzyme wieder und es wird spekuliert, dass durch „evolutive Flickschusterei“ (hier als „Kooption“ bezeichnet) diese Proteine schließlich zusätzlich auch als Linsencrystalline verwendet wurden. Hier stellt sich jedoch die Frage, welche verborgenen „Vormuster“ (pre-pattern) einer Linse zugrunde gelegen haben sollen. Eine Augenlinse entsteht ja nicht einfach durch eine Anhäufung eines gleichen (schon anderweitig verwendeten) Proteins, sondern bedarf weiterer komplexer Regulationen. Die Augenlinse ist z. B. eines der wenigen nicht durchbluteten Gewebe und hat damit eine Eigenschaft, die reguliert werden muss. Blutgefäße in der Linse, was als Krankheit auftreten kann, führen zur Trübung der Linse und damit zur Blindheit. Auch ist die Anordnung der Linsenzellen und ihrer enthaltenen Linsencrystalline nicht beliebig, usw.
Leider ist die Genetik der Augen bisher noch kaum verstanden, so dass über diese Spekulation der Kooption von Linsencrystallin eigentlich nicht geurteilt werden kann. Es scheint aber doch klar zu sein, dass ein einfacher „Kooptions“-Vorgang wie bei Insektenflügeln beim Auge nicht abgelaufen sein kann. Weiterhin müssen Evolutionsstadien durch selektionspositive Zwischenstufen miteinander verbunden sein. Dies ist bei den Flügelmustern denkbar (wenn auch unverstanden), denn diese Muster spielen im Brautwerbetanz der Fruchtfliegen eine Rolle. Durch Bevorzugung der Männchen mit jeweils den dunkelsten Flügelspitzen könnten hier solche selektionspositiven Übergänge postuliert werden. Letztlich bleiben aber auch diese Überlegungen Spekulation. Selektionspositive Übergänge bei Augenlinsen sind wegen ihrer Komplexität erst recht noch viel mehr spekulativ.
|
|
An/Aus: Noch mehr Flügelflecken
 |
| Grosses Bild |
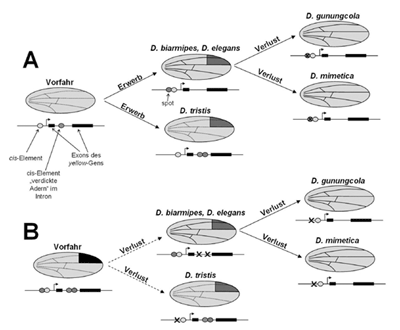
| Abb. 4: Vorschläge zum Evolutionsweg der Flügelflecken bei verschiedenen Fruchtfliegenarten. A: Vorschlag nach Prud´homme et al. (2006, umgezeichnet). Der vermutete Vorfahr der untersuchten Fruchtfliegenarten besitzt zwei cis-Elemente, die sich unabhängig voneinander so verändern, dass durch Kooption bestehender Stoffwechselwege Flügelflecken ausgeprägt werden (siehe Text bzw. Abb. 3). Nachfolgend kommt es bei einigen Arten durch den Verlust der entsprechenden regulatorischen Region auch wieder zum Verlust der Flügelflecken. B: Vorschlag mit einem komplexen Vorfahren, der alle Möglichkeiten der Flügelmusterung (Flecken, verdickte Adern) besaß. Durch Mutationen verändern sich die cis-Elemente, so dass Verluste in deren Regulationseigenschaften auftreten und dadurch auch die Flügelflecken bzw. die verdickten Adern ausfallen. Dieses Modell ist dadurch gedeckt, dass der Vorfahr aller untersuchten Fruchtfliegen immerhin mit einer Wahrscheinlichkeit von 28% ursprünglich Flügelflecken besaß. Verlustmutationen sind viel wahrscheinlicher und treten häufiger auf als Mutationen, die zum Erwerb neuer Eigenschaften führen (vgl. Winkler 2003). |
|
Wie bereits beschrieben, besitzen viele, aber bei weitem nicht alle Fruchtfliegen einen Flügelfleck. Innerhalb der sogenannten melanogaster-Gruppe gibt es 11 Arten mit und 8 Arten ohne Flügelfleck. Die Abstammungsverhältnisse dieser Fruchtfliegen-Arten sind bekannt und man beobachtet in dieser Gruppe sowohl den Verlust als auch vermutlich den unabhängigen (Neu-)Erwerb von Flügelflecken. Auf welche genetischen Änderungen sind die morphologisch sichtbaren Veränderungen zurückzuführen? In einer zweiten Arbeit untersuchte die gleiche Arbeitsgruppe die cis-Elemente des Gens yellow in verschiedenen Fliegenarten (Prud’homme et al. 2006). Die cis-Elemente wurden sequenziert, und in genetischen Experimenten wurde jeweils ihre Aktivität überprüft. Das Ergebnis zeigt, dass der Verlust des Flügelfleckens bereits nach einigen wenigen Punktmutationen in den cis-Elementen auftreten kann. Eine Tatsache, die nicht weiter verwunderlich ist, denn Mutationen treten ständig und zufällig auf, so dass auch für die Funktion wichtige Bereiche betroffen werden. Im Fall der nah verwandten Arten D. elegans (mit Fleck) und D. gunungcola (ohne Fleck) konnte gezeigt werden, dass mindestens zwei und höchstens sieben Punktmutationen im cis-Element spot (s. o.) für den Ausfall des Flügelflecks verantwortlich sind. Dies galt auch für eine weitere untersuchte Art ohne Flügelflecken (D. mimetica).
Da Mutationen ständig und ungerichtet auftreten, erscheint der Erwerb von Flügelflecken jedoch umso verwunderlicher. Denn nur wenn die einzelnen Mutationen positiv selektiert werden, kann ein Erwerb neuer Eigenschaften (hier von Flügelflecken) stattfinden (Weinreich et al. 2006). Die positive Selektion könnte mit den oben genannten Brautwerbetänzen der Männchen zusammenhängen, d.h. dass weibliche Fruchtfliegen in der Tat Männchen mit Flügelflecken (bzw. Ansätzen dazu) gegenüber solchen bevorzugen, die klare Flügel (ohne Flecken) haben. Allerdings bleiben diese Überlegungen bislang eine Spekulation.
Die Fruchtfliege D. tristis hat Flügelflecken und stammt aus der obscura-Gruppe von Fruchtfliegen, die acht Arten ohne Flecken und zwei Arten mit Flecken umfasst. Das entsprechende genetische cis-Element aus dieser Fruchtfliege wurde auf Flügelflecken-Aktivität hin getestet und interessanterweise war der Befund negativ. Das heißt: Ein anderes genetisches Element als in der melanogaster-Gruppe musste für Flügelflecken verantwortlich sein. Schließlich konnte dafür ein cis-Element im yellow-Gen selbst ausfindig gemacht werden (es war also in diesem Fall nicht dem Gen vorgeschaltet).
Die Gene der sogenannten höheren Organismen liegen nicht an einem Stück vor, sondern sind in Abschnitte unterteilt. Das fertige Protein entsteht nur, wenn die sinntragenden Abschnitte (Exons) aneinandergefügt werden, indem die Abschnitte ohne kodierende Funktion (Introns) ausgeschnitten werden. In einem solchen Intron befand sich ein weiteres cis-Element, welches die Produktion von yellow beeinflusste, intron y genannt. Außer im Flügelfleck wurde yellow in D. tristis auch besonders entlang der Flügeladern ausgeprägt, so dass diese besonders dunkel erscheinen. War nun dieses cis-Element im Intron von yellow in D. tristis neu entstanden oder lag lediglich eine Abänderung eines bereits vorhandenen cis-Elements vor? Um dies herauszufinden wurde D. guanche untersucht, eine weitere Art in der obscura-Gruppe ohne Flügelfleck. Tatsächlich fand sich auch bei dieser Art ein intron y cis-Element im Intron-Bereich des Gens yellow. Im Unterschied zu D. tristis verursacht dieses cis-Element aber keine Flügelflecken, sondern nur besonders dunkle Flügeladern. Damit, so die Autoren, entstanden die Flügelflecken auch bei D. tristis durch „Kooption“, d.h. die Veränderung eines bereits vorhandenen cis-Elements. Dieses cis-Element bindet ebenfalls bereits vorhandene trans-Faktoren und übernimmt dadurch deren Muster. Es handelt sich also auch nicht um eine komplette Neu-Entstehung (Abb. 4A).
|
|
Gleiche Wirkung – verschiedene Ursachen
Ein sehr interessanter Befund der durchgeführten Untersuchungen ist die Tatsache, dass zwei völlig verschiedene cis-Elemente, die an unterschiedlichen Orten im Genom mit unterschiedlicher Sequenz (spot und intron y) in zwei Fliegenarten jeweils sehr ähnliche Flügelflecken hervorrufen. Durch die Wirkung entsprechender trans-Elemente besteht ein räumliches Gerüst (Vormuster) im Flügel (Wray 2006). Es scheint so, als ob in Organismen genetische Module (hier: Ausbildung von Flecken) vorgegeben sind, die durch sehr unterschiedliche genetische Mechanismen („Ein-Schaltungen“) abgerufen werden können. Wie die zugrunde liegenden genetischen Module entstanden sind, wird durch solche Einsichten aber nicht erhellt.
|
Soll ein Protein von einem bestimmten Gen hergestellt werden, so muss erst eine Kopie („Blaupause“) vom Gen erstellt werden. Die Kopie („mRNA“) geht dann in die Protein-Produktion („Translation“). Je mehr solcher Gen-Kopien erstellt werden, um so mehr Protein kann produziert werden. Über diesen Schritt werden viele Gene reguliert, doch was legt die Kopienzahl fest? Gene lassen sich, grob vereinfacht, in zwei Abschnitte unterteilen: Eine Region, die für das herzustellende Protein selbst kodiert und eine davor liegende Region, die eine Steuerungsfunktion hat. Der Abschnitt mit der Steuerungsfunktion wird Promoter genannt. Der Promoter enthält bestimmte Abfolgen von DNA-Bausteinen, die von speziellen Proteinen, den Transkriptionsfaktoren, erkannt werden. Erst wenn ein solcher Transkriptionsfaktor an die DNA gebunden hat, kann in Folge das Enzym, welches für die Kopienherstellung notwendig ist, an die DNA andocken und eine Genkopie erstellen („Transkription“). Die Stärke der Bindung von Transkriptionsfaktoren an die DNA bestimmt über die Häufigkeit der Kopienherstellung: Bindet der Faktor gut, so können viele Kopien erstellt werden, bindet der Faktor schlecht, so kommt das Kopier-Enzym nur selten zum Zug. Mutationen im Promoter beeinflussen die Bindungsstärke des Transkriptionsfaktors und damit letztlich, wie häufig das Gen kopiert wird und dadurch wiederum, wieviel Protein hergestellt werden kann.
Die Sequenzabschnitte des Promoters, die die Transkriptionsfaktoren binden, werden als cis-Elemente bezeichnet (cis = diesseits), da sie unmittelbar vor (und manchmal auch in) dem Gen liegen. Die Transkriptionsfaktoren selbst werden als trans-Elemente bezeichnet (trans = jenseits), da sie von „irgendwo“ aus der Zelle kommen.
|
|
|
Erwerb oder Verlust?
Fasst man beide oben beschriebenen Publikationen über Flügelflecken bei Fruchtfliegen zusammen, so ergibt sich das folgende Bild: Die Fruchtfliegen-Art, welche der Vorfahr sowohl der melanogaster- als auch der obscura-Gruppe war, besaß einen kompletten Regulationsweg für das Protein Yellow, inklusive dazugehörender cis-Elemente. Zwei dieser cis-Elemente (spot und intron y) verändern sich durch Mutationen, so dass sie nun auch trans-Elemente binden können (und dadurch reguliert werden), die vorher bei der Fleckenbildung keine Rolle spielten. Durch die neue Regulation entstehen Flügelflecken. In beiden Fällen handelte es sich um die Abwandlung vorhandener cis-Elemente („Veränderung“) und nicht um eine komplette Neu-Entstehung („Neuheit“). Damit, so die Autoren, handelt es sich vermutlich wieder um Beispiele von „Kooption“: verborgene Muster verschiedener trans-Faktoren werden durch die veränderte Regulation des Proteins Yellow, basierend auf der Veränderung seiner cis-Elemente, sichtbar gemacht. Die durch die trans-Elemente bereits geschehende Regulation wird also von yellow übernommen.
Aber selbst dieser Befund ist meiner Ansicht nach nicht eindeutig. Die Autoren der zweiten Studie geben einen phylogenetischen Überblick über die Fruchtfliegen-Arten der melanogaster- und der obscura-Gruppe. Der letzte gemeinsame Vorfahr dieser insgesamt 29 Arten besaß mit einer Wahrscheinlichkeit von 28% bereits Flügelflecken! Es besteht daher durchaus die Möglichkeit, dass in keinem einzigen Fall ein Erwerb von Flügelflecken vorliegt, sondern immer nur ein Verlust, d.h. Mutationen veränderten die cis-Elemente dergestalt, dass die regulierenden trans-Elemente nicht mehr binden können (Abb. 4B). Die Datenbasis zur Klärung dieser Frage ist leider nicht breit genug, denn von den Autoren wurden nur beispielhaft die bereits genannten sechs Arten untersucht. Es wäre äußerst interessant, die genetischen Orte aller übrigen Arten dieser beiden Gruppen zu untersuchen.
Ein ähnlicher Fall wurde vor nicht allzu langer Zeit über die Flügel von Stabheuschrecken berichtet. Auch hier war man in einer ersten Publikation davon ausgegangen, dass einige Stabschrecken ihre Flügel verloren hatten, um sie dann – viele Generationen später – wieder zu erwerben. Nachfolgende kritische Betrachtungen kamen allerdings zum Schluss, dass es sich wohl immer nur um Verluste gehandelt hat. Der scheinbare Neuerwerb war ein Artefakt der phylogenetischen Rekonstruktion, denn man hatte dem Verlust wie auch dem Erwerb zunächst gleiche Wahrscheinlichkeiten zugeordnet. Ein Funktionsverlust ist aber sicherlich um Größenordnungen wahrscheinlicher (Winkler 2003).
Gehen wir davon aus, dass bei den Flügelflecken in der Tat doch eine „Kooption“ stattgefunden hat. Warum wäre das hier überhaupt möglich? Man weiß, dass Stoffwechselwege, die in einem Organismus nicht mehr gebraucht werden, durch weitergehende Mutationen innerhalb weniger Generationen so beschädigt werden, dass ein „Wieder-Einschalten“ nicht mehr möglich ist. Das heißt, der Ausfall dieser Stoffwechselwege hat – wenn er sowieso nicht mehr benötigt wird – keinen negativen Selektionswert; es ist quasi „egal“, ob dieser Weg existiert oder nicht. Dies wurde z.B. bei Blütenfarbstoffen der Windengattung Ipomea nachgewiesen. Die Blüten vieler Arten sind violett, weil ein roter und ein blauer Farbstoff hergestellt wird. In einer Art entfiel die Farbstoffsynthese für Blau. Dieser Syntheseweg wurde durch nachfolgende Mutationen so „zerschossen“, dass die Nachfahren dieser Art praktisch nie wieder zu einer violetten Blütenfarbe zurückkehren können, sondern bei Rot bleiben müssen (Winkler 2004).
Bei Yellow in der Fliege handelt es sich aber um einen besonderen Fall. Dieses Protein hat bei weitem nicht nur Einfluss auf die Farbgebung der Flügel, sondern auf viele andere Bereiche in der Entwicklung. Man spricht von einem pleiotropen Gen. Der Verlust eines solchen Gens ist extrem nachteilig, denn viele verschiedene Stoffwechselwege werden gleichzeitig beeinflusst, und das fein ausbalancierte System im Organismus gerät in eine Schieflage. Im Gegensatz zur relativ unwichtigen Blütenfarbe Blau bei der Windenpflanze ist also yellow in Fruchtfliegen ein wichtiges Gen. Das heißt, der Verlust von Flügelflecken führt nicht dazu, dass auf das dahinterliegende genetische System (welches u. a. Yellow umfasst) verzichtet werden kann, während der Verlust der blauen Blütenfarbe das dazugehörende genetische System dafür obsolet machte und schließlich völlig abschaltete. Der pleiotrope Charakter von yellow ist notwendige Voraussetzung, dass eine „Kooption“, d.h. ein Wieder-Einschalten der Flügelflecken überhaupt ablaufen kann. Ob es in der Tat so der Fall war, ist – wie oben bereits andiskutiert – nicht sicher entscheidbar.
Die Autoren der Fliegenstudie sehen in ihren Untersuchungen die von Jacob Monod postulierte „evolutionäre Flickschusterei“ bestätigt, bei der bestehende Elemente (hier z.B. yellow) in neue Funktionszusammenhänge eingebaut werden (vgl. Ullrich et al. 2006). Man muss aber in aller Deutlichkeit festhalten, dass es sich bei den aufgefundenen Mechanismen nur um Veränderungen in der Regulation eines Proteins (Yellow) handelt. Zudem ist eine echte „Kooption“, d.h. de novo Entstehung von Flügelflecken, m. E. nicht zweifelsfrei nachgewiesen worden.
Weiterhin ungeklärt bzw. nicht einmal im Ansatz verstanden sind Dinge wie die Entstehung des Melanin-Syntheseweges inklusive seiner Notwendigkeit für den Gesamtorganismus als „biochemische Maschine“, die Verhaltensänderungen beim Brautwerbetanz, die Entstehung des genetischen Steuerungssystems aus cis- und trans-Elementen, usw. Flügelflecken sind nur ein winziger und offensichtlich entbehrlicher Bestandteil eines komplexeren Ganzen. Veränderungen der Regulationen bestehender Systeme sind sicherlich Bestandteil einer Evolutionstheorie, aber nur der einer Mikroevolution. Die viel grundlegendere Frage nach der Entstehung des Gesamtsystems, also einer Makroevolution bleibt unbeantwortet. Der Schluss von den beschriebenen mikroevolutiven Prozessen auf Makroevolution, wie ihn die Autoren der Fliegenstudien bei der Evolution der Wirbeltierlinse ziehen, ist durch die Daten ihrer Studie nicht gedeckt. Denn Veränderungen in der Regulation von Genen sind nur eine notwendige Bedingung (von vielen weiteren!) für Evolution, aber keine hinreichende für deren Neuentstehung. Es liegt noch viel Forschungsarbeit vor uns, um diese Grenze mit mehr Klarheit zu erfassen.
|
Literatur
- Eckstein (1930)
- Der Birkenspanner (Amphidasis betularius). Journal of Pest Science 6, 59-60.
- Gompel N, Prud’homme B, Wittkopp PJ, Kassner VA & Carroll SB (2005)
- Chance caught on the wing: cis-regulatory evolution and the origin of pigment patterns in Drosophila. Nature 433, 481-487.
- Prud’homme B, Gompel N, Rokas A, Kassner VA, Williams TM, Yeh SD, True JR & Carroll SB (2006)
- Repeated morphological evolution through cis-regulatory changes in a pleiotropic gene. Nature 440, 1050-1053.
- Ullrich H, Winkler N & Junker R (2006)
- Zankapfel Auge. Ein Paradebeispiel für „Intelligent Design“ in der Kritik. Studium Integrale Journal 13, 3-14.
- Weinreich DM, Delaney NF, Depristo MA & Hartl DL (2006)
- Darwinian evolution can follow only very few mutational paths to fitter proteins. Science 312, 111-114.
- Winkler N (2003)
- Drei Schritte vor – und zwei zurück: Flügel bei Stabschrecken. Studium Integrale Journal 10, 81-83.
- Winkler N (2004)
- Einschränkung durch Evolution? Studium Integrale Journal 11, 80-82.
- Wray GA (2006)
- Evolution: spot on (and off). Nature 440, 1001-1002.
|
|  |
