|
|
|
|
|
Hypothesen über primitive und fortschrittliche AngiospermenmerkmaleWie im ersten Teil des Aufsatzes erwähnt, befassen sich - mangels überzeugender Fossilüberlieferung - die meisten Lehrbuchdarstellungen über die Abstammung der Angiospermen hauptsächlich mit der Frage, wie die ersten Angiospermen ausgesehen haben könnten, und nur ganz am Rande mit der Frage, wie sie wirklich ausgesehen haben. Das heißt, es ist in erster Linie die subjektive Einschätzung des Autors, was als primitiv und was als fortschrittlich anzusehen ist. Dieser Tatbestand wird z.B. bei Raven et al. (2000, S. 555) korrekt wiedergegeben, allerdings ohne daß daraus Konsequenzen gezogen würden: "All unsere Vermutungen über die Evolution der Blüte beruhen auf vergleichenden Studien heutiger Formen und aus dem, was wir aus Fossilfunden erschließen können." Bestimmte Vorstellungen haben eine besondere Popularität erlangt, und so wird meist folgende Vorstellung von den Urangiospermen vermittelt: Kleinere Holzpflanzen mit ungeteilten, fiedernervigen Blättern. Blüten relativ groß (etwa nach Art der Magnolien) mit spiraliger Anordnung der jeweils zahlreich vorhandenen Blütenglieder. Blüten zwittrig mit Anpassungen an Insektenbestäubung, evtl. durch Käfer. Blütenhülle einfach, d.h. Kelch und Krone noch nicht geschieden. Fruchtblätter noch nicht verwachsen mit jeweils zahlreichen Samenanlagen, zur Fruchtreife balgartig. Zur Demonstration fällt es nicht schwer, rezente oder ausgestorbene Vertreter zu finden, die mindestens einige dieser Merkmale zeigen. Besonders augenfällig ist das etwa in Raven et al. (2000, S. 551ff). Die Autoren greifen die im ersten Teil des Aufsatzes erwähnte fossile Gattung Liriophyllum (= Archaeanthus) heraus mit ihren großen Blüten, in denen an einer langen Achse in spiraliger Anordnung mehr als 50 vielsamige Fruchtblätter sitzen. Solche Angaben haben prägende Wirkung auf den Leser, selbst wenn an anderer Stelle (1988, S. 410) einschränkend angemerkt wird: "Die langgestreckte, zapfenförmige Gestalt des Magnoliaceen-Receptaculums ist sicherlich eine Spezialisierung, die von anderen primitiven Angiospermen-Familien nicht geteilt wird."
Was aus den Lehrbüchern kaum hervorgeht, ist die Tatsache, daß es zu allen Zeiten konkurrierende Auffassungen gegeben hat, die in der Sekundär- und Primärliteratur mehr oder weniger heftig diskutiert werden. Es kann daher nicht oft genug betont werden, wie sehr subjektiv oft die Einschätzungen sind, ob etwas als ursprünglich oder abgeleitet zu gelten hat. Tab. 4 zeigt am Beispiel der alpinen Krautweiden auf, daß das Merkmal der Windblütigkeit (= Bestäubung der Blüten durch den Wind) nach gängigen Vorstellungen immer wieder im Laufe der Phylogenie zwischen primär und sekundär hätte wechseln müssen. Tab. 5 stellt verschiedene Auffassungen über primitive und fortschrittliche Merkmale bei Angiospermen zusammen. Fett gedruckt sind gängige Auffassungen, kursiv gedruckt Angaben über Zustände, die für sekundär gehalten werden. |
|
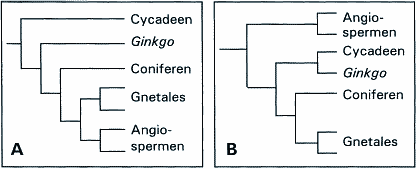
In die Diskussion über das Erscheinungsbild der primitiven Angiospermen ist in den letzten 5-10 Jahren wieder Bewegung gekommen, vor allem weil - wie oben angesprochen - neuere Fossilfunde nicht mit den theoretischen Erwartungen übereinstimmten. Zum Teil vollzieht sich daher eine Angleichung der theoretischen Erwartung an die reale Situation. Man vergleiche dazu etwa, daß Strasburger von der vorletzten zur letzten Auflage (1991, 1998) die Ansicht über die ursprüngliche Blütengröße (kommentarlos) verändert hat (während er allerdings noch an der holzigen Wuchsform als ursprünglich festhält). Taylor & Hickey (1996, S. 3) fassen den gegenwärtigen Stand der Diskussion zusammen. Nach ihrer Ansicht sind von den verschiedenen im Laufe der Zeit vorgeschlagenen Hypothesen zur Situation der Urangiospermen nur noch die beiden folgenden ernsthaft in Erwägung zu ziehen. Sie selber sowie u.a. auch Endress (1993) tendieren eindeutig zur zweiten Hypothese: 1. Die "Magnoliales"-Hypothese: Nach dieser klassischen Vorstellung waren die ersten Angiospermen kleine baumförmige Holzpflanzen mit großen, vielgliedrigen Blüten. Als Ausgangsgruppe stellt man sich etwa Formen mit Fortpflanzungsorganen wie die der Bennettiteen vor. 2. Die "Herbaceous Origin-Hypothese": Diese Hypothese stellt eine Variante der Paleoherb-Hypothese von Donoghue & Doyle (1989) dar, wonach die ersten Angiospermen in jenen Ordnungen der Magnoliopsida zu suchen sind, die überwiegend krautige Vertreter haben, also z. B. Nymphaealus, Chloranthales und Piperales. Demnach waren die ersten Angiospermen nicht verholzte Pflanzen, sondern krautige, ausdauernde Rhizompflanzen. Außerdem besaßen sie zahlreiche, kleine, weniggliedrige Blüten, hatten also nicht jene großen und vielgliedrigen Blüten, wie man sie sich nach der Magnoliales-Hypothese herkömmlicherweise als typisch für ursprpüngliche, durch Insektenbestäubung gekennzeichnete Angiospermenblüten vorstellte. Sind die Angiospermen monophyletisch?Die Frage, ob Angiospermen eine natürliche Abstammungseinheit darstellen, d.h. einheitlich (monophyletisch) oder heterogen (polyphyletisch) sind, ist für die Beurteilung ihrer Abstammung wesentlich. Im ersteren Fall würden sie sich von einer der vorhergehenden Gruppen ableiten, im zweiten Fall wären sie konvergent aus verschiedenen Vorfahren entstanden. Wegen fehlender Zwischenstufen ist man auch hier auf theoretische Überlegungen und auf den Vergleich rezenter Formen angewiesen. Die überwiegende Zahl der Autoren betrachtet heute die Angiospermen als monophyletische Gruppe. Crane et al. (1995) halten aufgrund neuerer kladistischer Analysen die Angiospermen sogar für "the most strongly supported monophyletic group in the plant kingdom". Strasburger (1998) nennt als Argumente für Monophylie: 1. Die Angiospermen sind durch zahlreiche Merkmale verbunden, d.h. sie haben untereinander sehr viele Gemeinsamkeiten. 2. Es besteht gegenüber allen anderen Samenpflanzen eine sehr deutliche Formenlücke, während innerhalb der Angiospermen nirgends derart unüberbrückbare Formenlücken gefunden werden können. Es gibt aber auch gegenteilige Stimmen. So spricht sich Hughes (1994) für Polyphylie aus und Carlquist (1996) zeigt am Beispiel der Holzanatomie, wie hochgradig polyphyletisch die Angiospermen in diesem Bereich sind. Kladistische Verfahren zur Abstammung der AngiospermenIn letzter Zeit werden zunehmend kladistische Verfahren zur Aufhellung der Systematik und Abstammung der Angiospermen eingesetzt. Man versucht also, aus dem Vergleich von überwiegend oder ausschließlich rezenten Formen auf die Vergangenheit rückzuschließen. Daß entsprechende Kladogramme je nach betrachteten Merkmalen oftmals nicht in Deckung gebracht werden können, überrascht nach den Erfahrungen bei anderen Organismen kaum. Abb. 9 zeigt dies anschaulich am Beispiel des Vergleiches morphologischer und molekularer Charakteristika (hier ITS-Sequenzen von Chloroplasten-DNA). Betrachtet man etwa die Gnetales (Gnetum und Verwandte, vgl. den 1. Teil dieses Aufsatzes), so stehen diese nach der Morphologie den Angiospermen, nach den genannten DNA-Sequenzen aber den übrigen Gymnospermen näher. |
|
|
Trotz des Vorteils morphologischer Vergleiche, die mindestens zum Teil auch fossiles Material berücksichtigen können, spielen heute sog. "molekulare Stammbäume" eine absolut dominierende Rolle. Eine aktuelle Zusammenfassung findet sich bei Sytsma & Baum (1996), vgl. auch Frohlich & Parker (2000). Die Probleme bei der Interpretation solcher Verfahren sind sehr vielfältig (vgl. etwa Sytsma & Baum 1996). Es hier nicht der Platz, auf die Problematik im Einzelnen einzugehen. Wohl kann zusammenfassend gesagt werden, daß alle Ergebnisse und Interpretationen mit großer Sorgfalt bzw. Vorsicht betrachtet werden müssen und daß bei unterschiedlichen Methoden u.U. stark widersprüchliche Ergebnisse zustande kommen können. Diese Problematik wird auch offen diskutiert. Offensichtlich besteht aber ein solch großes allgemeines Interesse an entsprechenden Ergebnissen, daß die vorläufigen Dendrogramme immer wieder Beachtung finden und in einer Art und Weise präsentiert werden, als handle es sich um gesicherte Fakten. So hat z.B. eine entsprechende Graphik auch in das Lehrbuch von Strasburger (1998) Eingang gefunden. Auf einige Probleme sei kurz hingewiesen: 1. Die Wahl der Außengruppe kann das Aussehen des Kladogramms stark beeinflussen. 2. Nicht selten liegen für unterschiedliche Taxa unterschiedliche Basenaustauschraten vor, was die Zusammenführung der Daten sehr erschwert (vgl. Taylor & Hickey 1996). 3. Je länger die mutmaßlichen Gabelungen zurückliegen, desto größer wird die Fehlerquelle. Diese ist in unserem Fall besonders hoch, da es ja um die allerersten Anfänge der Angiospermenentwicklung geht. Einer der ersten Versuche zur Erstellung eines Stammbaums der Angiospermen aufgrund des Vergleichs von DNA-Sequenzen stammt von Martin et al. (1989). In dem Fall ging es um die GAPDH (Glycerinaldehydphosphatdehydrogenase). Folgt man den Autoren, hätte die Trennung der Monokotylen und Dikotylen zu dem überraschend frühen Zeitpunkt von etwa 319 Millionen Jahren vor heute stattgefunden, also irgendwann im Karbon. Diese Interpretation wurde heftig angegriffen, u.a. mit der Begründung, das zugrunde liegende Datenmaterial wäre zu gering und paläobotanische Befunde wären zu wenig berücksichtigt worden (vgl. u.a. Sytsma & Baum 1996). Seither wurden zahlreiche "molekulare Stammbäume" vorgelegt. Sie sind sehr verschieden, doch scheint sich für den Abzweig der Angiospermen von ihren mutmaßlichen Vorfahren eine gewisse Häufung im Bereich der Trias abzuzeichnen (Taylor & Hickey 1996). An dieser Stelle soll kurz auf die aktuelle Diskussion über die ursprünglichsten Angiospermen eingegangen werden (vgl. Crepet 2000). In den Jahren 1998 und 1999 erschienen mehrere Arbeiten, die unabhängig voneinander vornehmlich aufgrund des Vergleiches der DNA von Regulationsgenen (MADS-Box-Genen) zu dem Ergebnis kamen, daß der Strauch Amborella trichopoda, ein Endemit der im Südpazifik gelegenen Insel Neukaledonien, weit isoliert von allen anderen Bedecktsamern steht. Da die Gattung als Vertreter der Ordnung Chloranthales zu den "Paleoherbs" gehört, also zu jenen Magnolienverwandten, die u.a. wegen gewisser Fossilfunde von einigen Autoren als älteste Angiospermen angesehen werden, war die Antwort schnell parat und wurde so auch in der New York Times verkündet: "Hier haben wir den primitivsten Bedecktsamer vor uns." Entsprechend groß war ein plötzliches Interesse vieler Botaniker an dieser Pflanze, die im Übrigen gar nicht so urtümlich erschien. Inzwischen wurde die mutmaßliche Rolle von Amborella aber schon wieder relativiert (vgl. z. B. Barkman et al. 2000, Graham et al. 2000), indem sich diese Gattung den "untersten Ast" im "Stammbaum" der Angiospermen z.B. mit der Seerose Nymphaea teilen muß. Damit tauchen jetzt wieder andere Probleme der Interpretation auf, da die beiden Gattungen bezüglich zweier für Merkmalsabfolgen bedeutender Merkmale total verschieden sind: Amborella besitzt keine Gefäße (Tracheen) und ist eingeschlechtig, während Nymphaea Tracheen und zwittrige Blüten besitzt. Interessant sind bei dieser Diskussion zwei Dinge, zum einen die starke Abhängigkeit der Ergebnisse von Methodenwahl und Interpretation und zum anderen die zwischen den Zeilen zu lesende Vorstellung, die Frage der Abstammung der Angiospermen habe im Wesentlichen damit zu tun, welche rezente Gattungen aufgrund molekularer Vergleiche als primitiv gewertet werden könnte und welche nicht. Wir hatten im ersten Teil dieses Artikels schon angesprochen, daß die Situation der starken Radiation der Angiospermen zu Beginn der Kreide und das Fehlen angiospermer Fossilien vor der Kreide nach einer Erklärung verlangt, wo und wie und wie lange die zu erwartenden Zwischenformen existiert haben könnten. Wenn nun aufgrund der DNA-Vergleiche die Urangiospermen zeitlich in die Trias oder gar noch früher zurückverlegt werden, bleibt natürlich die Frage, wieso die Angiospermen über etwa 100 Millionen Jahre (oder über einen noch längeren Zeitraum) hinweg existieren konnten, ohne daß sie Fossilbelege hinterließen. Welche Pflanzengruppe kommt als Vorfahre der Angiospermen in Betracht?Wegen der fehlenden Zwischenglieder ist diese Frage schwer zu beantworten. Während also das "abominable mystery" unverändert im Raum steht, gibt es zahlreiche hypothetische Vorstellungen zur Lösung des Rätsels. Alle Versuche haben damit zu kämpfen, daß die in Frage kommenden Gruppen, und zwar sowohl was die rezenten als auch was die fossilen Vertreter angeht, in vielerlei Hinsicht einschließlich Blütenbereich stark spezialisiert sind. Das ist auch weitgehend unabhängig davon, wann man eine Abzweigung der Angiospermen von ihrer Schwestergruppe annimmt. Allerdings lehnen die meisten Autoren eine Ableitung der Angiospermen von fossil erhaltenen Pflanzenarten der Kreide ab. Da die Nacktsamer lange vor den Bedecktsamern fossil erhalten sind, gehen nahezu alle Theorien davon aus, daß in diesem Bereich die Vorfahren der Angiospermen zu erwarten sind. Lediglich im Zusammenhang mit jenen molekularbiologischen Daten, die eine sehr frühe Entstehung der Angiospermen nahelegen, wurde vermutet, daß Angiospermen und Gymnospermen eine gemeinsame Wurzel im Bereich der Farnartigen haben könnten. Im Laufe der Zeit sind die meisten Hauptgruppen der Gymnospermen als Vorfahren der Angiospermen in Erwägung gezogen worden (Stewart & Rothwell 1993). Zwar finden sich hier und da einzelne Merkmalskomplexe, die im Sinne von Vorstufen gedeutet werden können (z.B. weibliche Blütenorgane bei bestimmten Taxa), aber für Merkmalsabfolgen im Sinne einer Abstammung müßten die beiden folgenden Kriterien erfüllt sein, was aber praktisch nie der Fall ist. 1. Die Abfolge muß - etwa ausgehend von farnartigen Pflanzen über gegebenenfalls mehrere Stufen nacktsamiger Pflanzen bis hin zu heutigen bedecktsamigen Pflanzen - tatsächlich oder doch mindestens theoretisch gezeigt werden können. Nach Auffassung von Schweitzer (1989) ist der Weg, der zu den männlichen Organen der Angiospermen führt, ausgehend von den Mikrosporangien verschiedensporiger Farne, durch zahlreiche Übergangsformen theoretisch belegt. Nach dem gleichen Autor ist jedoch die Abstammungsfrage der weiblichen Organe völlig unklar, so daß hier nicht einmal überzeugende theoretische Überlegungen vorgelegt worden sind. Im gleichen Sinne schreibt Strasburger (1998), die morphologische Interpretation der Karpelle (= Fruchtblätter) sei unklar. 2. Es reicht nicht aus, wenn Abstammungsreihen für einen ausgewählten Merkmalskomplex gezeigt werden, vielmehr muß die Abfolge auch in anderen Bereichen stimmen. So nützen z.B. Reihen fossiler Belege für die Entwicklung von männlichen Blütenteilen nichts, wenn nicht an denselben Belegen auch die Entwicklung der weiblichen Blütenteile gezeigt werden kann bzw. sich dort eine andere Abfolge nahelegt. Dabei ist es allerdings nicht nötig, daß alle Organe die gleiche Entwicklungsgeschwindigkeit aufweisen. Die Coniferophytina (vgl. Abb. 7 im 1. Teil) werden trotz ihres hohen Alters im Allgemeinen als Vorfahren ausgeschlossen (vgl. Strasburger 1998), da sie insgesamt sehr spezialisiert sind. Dies gilt etwa für die hier besonders interessierenden männlichen und weiblichen Blüten sowie für die Nadelblätter. Es bleiben also nur die Cycadophytina (vgl. Abb. 8 im 1. Teil) übrig, bei denen vor allem die fiedrige Grundstruktur der Blätter und verschiedene Blütenmerkmale als Anklänge an die Angiospermen aufgefaßt werden können. |
|
|
Die Gnetopsida mit ihren zum Teil zwittrigen Blüten, Ansätzen zur Tierbestäubung, teilweise stark reduzieten Gametophyten und doppelter Befruchtung wurden schon frühzeitig als Vorläufer der Angiospermen diskutiert. Nach der klassischen Pseudanthientheorie (Abb. 10) stellte man sich vor, daß die Angiospermenblüte aus einem Blütenstand mit benachbarten geschlechtsverschiedenen Blüten hervorgegangen ist, etwa vergleichbar mit der Situation der Wolfsmilch-Arten (Euphorbia), wo jeder Teilblütenstand mit jeweils mehreren männlichen und einer weiblichen Blüte wie eine zwittrige Einzelblüte erscheint. Die Staubblätter wären also aus männlichen Blüten, die Fruchtblätter aus weiblichen Blüten entstanden. Diese Theorie hat über längere Zeit kaum Anhänger gefunden. Seitdem aber nicht nur wegen morphologischer, sondern auch wegen bestimmter molekularbiologischer Daten die Gnetopsida von einigen Autoren als Schwestergruppe der Angiospermen angesehen werden (vgl. Taylor & Hickey 1996, Strasburger 1998) ist die Pseudanthientheorie wieder neu im Gespräch. Trotz allem sind sich die meisten Autoren darüber im Klaren, daß die drei Gattungen Gnetum, Ephedra und Welwitschia als stark abgeleitet gelten müssen, und der Interpretation als Basisgruppe die Tatsache entgegensteht, daß sie fossil praktisch nicht dokumentiert ist. Daher neigt man heute eher dazu, hier eine Parallelentwicklung zu sehen. Strasburger (1998) formuliert es so: "Einer Ableitung von den Gnetopsiden steht aber die Schwierigkeit entgegen, die Angiospermenblüte als Blütenstand zu interpretieren. Auch ist es wenig wahrscheinlich, daß so stark reduzierte rezente Restgruppen wie die heutigen Gnetopsida am Anfang der offenbar sehr alten Bedecktsamer stehen sollten." Im Übrigen zeigt sich auch an dieser Stelle, wie vorsichtig man mit der Interpretation molekularbiologischer Daten sein muß. Denn während nach Taylor & Hickey (1996) angeblich Übereinstimmung darin besteht, daß die Gnetopsida die nächst verwandte rezente Gruppe der Angiospermen darstellen, stehen nach jüngsten molekularen Untersuchungen (vgl. Crepet 1998; 2000) die Gnetopsida den übrigen Gymnospermen näher als den Angiospermen. Im gleichen Sinne kommen Qiu et al. (1999) nach Vergleich von DNA-Abschnitten von Mitochondrien, Plastiden und Zellkern aus 63 Familien zu dem Schluß "that Gnetales are related to the conifers and are not sister to the angiosperms, thus refuting the Anthophyte Hypothesis". Das heißt, die Autoren sehen schwerwiegende Einwände gegen die Vorstellung, die Vorläufer der Angiospermen in der Verwandtschaft von Gnetum oder bei den im Folgenden angesprochenen Bennettiteen zu suchen, die wegen ihrer besonders blütenähnlichen Fortpflanzungsorgane auch als frühe Anthophyten bezeichnet werden. Die ausgestorbenen Bennettitopsida, die oft als Vorläufer der Gnetopsida angesehen werden, zeigen in ihren Blüten und hier wiederum vor allem durch die Staubblätter und die Blütenhülle gewisse Anklänge an die Angiospermen. Doch lassen sich nur die Staubblätter und die Blütenhülle zwanglos als Vorläufer interpretieren, bei den eigenartigen Fruchtblättern ist dies kaum möglich. Nach Ansicht von (Taylor & Hickey 1996) sind aufgrund von DNA-Vergleichen die Bennettitopsida eindeutig die nächst verwandte ausgestorbene Gruppe. Man vergleiche dazu aber die im vorigen Abschnitt zitierten Aussagen. Bei den Cycadopsida (Palmfarne) bietet es sich an, die flächigen Fruchtblätter (Megasporophylle) mit ihren randständigen Samenanlagen als Vorläufer der Angiospermenfruchtblätter zu verstehen. Dies findet seinen Ausdruck in der Euanthientheorie (Abb. 10). Danach hatten die Urangiospermen ebenso wie ihre heutigen Vertreter echte Blüten, also Euanthien. Die Fruchtblätter entstanden wie geschildert aus Megasporophyllen und die Staubblätter sowie die Blütenhülle (Perianth) gehen auf vergleichbare Gebilde der Bennettiteen zurück. Hier ergeben sich aber zwei Probleme: 1. Blütenorgane derselben Pflanze können sich schlecht von zwei verschiedenen Taxa ableiten. 2. Cycadeen und Bennettiteen sind in anatomisch-morphologischer Hinsicht zu abweichend, als daß sie problemlos als die gesuchte Ausgangsgruppe angesehen werden könnten. So bleibt noch der Versuch der Ableitung von den längst ausgestorbenen älteren Samenfarnen (Lyginopteridopsida) oder vielleicht sogar von deren Vorfahren, womit die Entstehung der Angiospermen vermutlich nicht nur um etwa Hundert Millionen Jahre, sondern sogar mehrere Hundert Millionen Jahre früher angesetzt werden müßte als der Beginn ihrer explosiven Radiation. Auf jeden Fall stehen die Samenfarne den Angiospermen nicht sehr nahe, so daß es kaum ausführliche Diskussionen über eine entsprechende Abstammungsfolge gibt. Allerdings werden (wie im 1. Teil schon erwähnt) aufgrund von molekularbiologischen Vergleichen (vgl. Frohlich & Parker 2000, Crepet 2000) die Samenfarne neu als potentielle Vorfahren der Angiospermen ins Gespräch gebracht. |
Das plötzliche, ja geradezu explosionsartige Auftreten der Angiospermen in der Kreide verbunden mit jeglichem Fehlen überzeugender fossiler Belege vor der Kreide bleibt eines der großen Geheimnisse in der Abstammung der Organismen. Zwar hat man in den letzten Jahren eine Reihe sehr früher Funde von Angiospermen tätigen können, doch waren diese oft ziemlich spezialisiert bzw. trugen Merkmalskombinationen, die keine Brücke zu irgendwelchen potentiellen Ausgangsgruppen hätten schlagen können. Hinzu kommt, daß neuere Pollenuntersuchungen bisherigeVorstellungen einer Abfolge von primitiven Einfurchen-Pollenkörnern der Farnpflanzen und Nacktsamer bis hin zu den fortgeschrittenen Dreifurchen-Typen moderner Bedecktsamer zerschlagen haben. Dies alles macht neues Nachdenken erforderlich, und liebgewordene Vorstellungen über die primitiven Angiospermen müssen zum Teil aufgegeben werden, da theoretische Überlegungen und konkrete Funde nicht in Deckung gebracht werden können. Selbst so grundlegend erscheinende Fragen wie die, ob die ersten Angiospermen krautig oder holzig waren, sind alles andere als geklärt. Die alte Frage steht unverändert im Raum: Wo waren die in reicher Zahl zu erwartenden Zwischenstufen zwischen Gymnospermen und Angiospermen vor der Kreide über einen Zeitraum von + 100 Millionen Jahren (gängiger Zeitrechnung)? Wieso haben sie keinerlei Spur hinterlassen? Diese Frage bleibt auch, wenn man, wie zunehmend üblich, durch Vergleich von DNA-Sequenzen rezenter Formen per Extrapolation auf das Alter der frühesten Angiospermen und auf ihre mögliche Ausgangsgruppe zurückrechnet. Außerdem zeichnet sich schon jetzt ab, daß die Situation durch Hinzunahme weiterer Sequenzen komplizierter statt einfacher wird - ein Phänomen, das allgemein für "DNA-Stammbäume" zu gelten scheint. Wie dem auch sei, Vergleiche rezenter Formen werden denjenigen kaum überzeugen, der nach klassischem Ansatz phylogenetischer Forschung der konkreten Merkmalsabfolge von angenommenen Vorläufern bis hin zu den rezenten Formen nachgeht. An dieser Stelle sind wir aber praktisch nicht weitergekommen. Selbst so alte Fragen wie die, ob die Zwitterblüte der Angiospermen auf eine Einzelblüte oder einen Blütenstand mit eingeschlechtigen Blüten zurückgeht, wird nach wie vor kontrovers diskutiert. Sollten etwa - wie in jüngster Zeit wieder neu in die Diskussion geworfen - Samenfarne den Ausgang der Angiospermen bilden, so bleibt nicht nur die Frage der Merkmalsfolge offen, sondern es rückt auch der zu überbrückende Zeitraum zwischen Entstehung und Fossilüberlieferung in einen Zeitrahmen von vielleicht 200 Millionen Jahren oder mehr (nach gängiger Datierung). Es scheint, daß wir noch eine Zeitlang mit dem "abominable mistery" leben müssen, selbst wenn einige Zeitgenossen die Lösung des Problems unmittelbar bevorstehen sehen. |
Literatur
|

|