|
|
|
|
|
Zur Abstammung der Angiospermen, der bedecktsamigen Blütenpflanzen, liegt zwar eine umfangreiche Literatur vor, aber nur ein kleiner Teil der Publikationen befaßt sich mit fossil erhaltenen Resten von Angiospermen. Der überwiegende Teil der Literatur widmet sich theoretischen Überlegungen zur Frage, wie man sich die ursprünglichen Angiospermen vorzustellen hat. Diese Überlegungen basieren weitestgehend auf dem Vergleich heute lebender Formen: "The literature available on the angiosperm origin is voluminous, but is concerned in the main with extant forms" (Meyen 1987, S. 231). Das hat damit zu tun, daß der Fossilbericht bisher wenig Befriedigendes zur Frage der Abstammung der Angiospermen beigetragen hat. "So eindrucksvoll die Aussagen der Paläobotanik hinsichtlich der zeitlichen Entfaltung der Angiospermen auch sind, zur Frage der Herkunft und Stammesgeschichte dieser Gruppe tragen sie bislang kaum bei" (Strasburger 1998). Bekanntlich war für Charles Darwin das schlagartige Auftreten der Angiospermen (in der Kreide) und das völlige Fehlen von Zwischenformen zu anderen Pflanzengruppen ein großes Problem, ein "abominable mystery" (abscheuliches Rätsel oder Geheimnis; vgl. F. Darwin & Seward 1903). Selbstkritisch schrieb er dazu, daß dies wohl einer der stärksten Einwände gegen seine Theorie wäre. Er rechnete aber fest damit, daß weitere Fossilfunde das Problem lösen würden.
Zwar sind Fossilien der Angiospermen seit Darwin in reichem Maße gefunden worden, aber der Lösung des Problems ist man praktisch nicht näher gekommen. Darüber sind sich die meisten Autoren einig (vgl. Stewart & Rothwell 1993). Strasburger (1998) faßt die Situation wie folgt zusammen: "Die stammesgeschichtliche Herkunft der Angiospermen ist noch ungeklärt." Dieser Satz ist von den vorhergehenden Auflagen des Lehrbuches unverändert übernommen worden. Die Einschätzung der in den letzten Jahren erzielten Fortschritte ist unterschiedlich. So schreiben Taylor & Hickey (1996, S. 7): "Rapid progress is being made. The extent of the "abominable mystery" of angiosperm evolution is rapidly diminishing and the pace of research is now such that we feel that, in 20 years, the issue will be no more mysterious than for any other major plant group." Hier wäre allerdings kritisch anzumerken, daß auch bei allen anderen "größeren Pflanzengruppen" die Abstammungsverhältnisse keineswegs klar sind (vgl. etwa Junker 1996 für die Abstammung der frühen Landpflanzen). - Crepet (1998) ist weit weniger optimistisch: "There is so much uncertainty about angiosperm relationships that bona fide evidence of very early angiosperms should be greeted with an enthusiasm fueled by desperation." Nach der ernüchternden Darstellung zahlreicher Fakten, die die Suche nach der Angiospermen-Herkunft erheblich erschweren, überrascht allerdings die vom gleichen Autor wenig später gegebene Einschätzung: "I predict that the great "abominable mystery", with us for over 100 years, will not last another 10." In den letzten Jahren hat man einige interessante und auch in der Tagespresse gewürdigte Fossilfunde der "ältesten" Angiospermen tätigen können. Aber auch diese Funde haben nicht zur Lösung des "Rätsels" beigetragen, weil sie mehrheitlich nicht den aufgrund theoretischer Überlegungen erwarteten Vorstellungen entsprachen (z.B. krautige Pflanzen mit kleinen Blüten anstelle der erwarteten Holzpflanzen mit großen, vielgliedrigen Blüten), und weil die bisherigen Funde wegen ihrer hohen Spezialisierung nicht als evolutionäre Übergangsformen gewertet werden können. Die dadurch veränderte Situation hat noch kaum Eingang in die Lehrbücher gefunden. Immerhin ist z.B. im Strasburger (1998), dem bekannten Botanik-Lehrbuch, zu erkennen, daß die Charakterisierung der primitivsten Angiospermen als Holzpflanzen mit großen mehrgliedrigen Blüten nicht mehr als alleinige Sichtweise gilt. |
Die Angiospermen heuteDie Angiospermen umfassen heute etwa 250 000 Arten in etwa 450 Familien und sind damit die bei weitem artenreichste Gruppe des Pflanzenreichs (vgl. Tab. 1). Ihre Artenzahl ist höher als die aller anderen grünen Pflanzen zusammen. Das Spektrum der Variabilität ist gewaltig. Es reicht z.B.von den winzigen Zwergwasserlinsen (Wolffia) von etwa 1 mm Durchmesser bis zu über 100 m hohen Eucalyptusbäumen oder bis über 200 m langen Lianen. Ökologische Anpassungen gibt es sehr ausgeprägt, z.B. bei Pflanzen der Wüsten, des Hochgebirges, der Seen und Flüsse, bei Epiphyten (Aufsitzer) der Regenwälder oder bei Schmarotzerpflanzen, um nur einige zu nennen. |
Die Angiospermen wurden lange Zeit in die beiden bekannten Klassen der Dikotylen (Zweikeimblättrige) und Monokotylen (Einkeimblättrige) eingeteilt. Neuerdings wird aufgrund kladistischer inkl. molekularbiologischer Analysen eine Dreiteilung vorgeschlagen, indem die Dikotylen in zwei eigenständige Klassen aufgeteilt werden. Dabei stellt man sich die Stammesgeschichte so vor, daß von ursprünglichen Dikotylen ausgehend, die man jetzt als Magnoliopsida s.str. abtrennt, sich einerseits die übrigen "fortschrittlichen" Dikotylen (= Eudicots oder Rosopsida), andererseits die Monokotylen (= Liliopsida) abgeleitet haben (Tab. 1, Abb. 6). Fossile Pollenkörner der Angiospermen
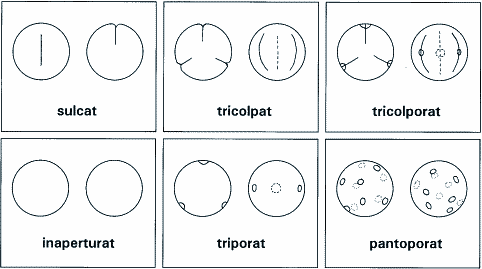
Die ältesten sicher den Angiospermen zuzuordnenden fossilen Pflanzenreste sind Pollenkörner. Dies ist eine deutliche Parallele zu den Sporen der devonischen Landpflanzen, die ebenfalls der Überlieferung von Makrofossilien weit vorauseilen. Pollenfunde reichen bis in die Kreide-Serie des Barremiums (vgl. Tab. 2), zum Teil sogar bis in die des Hauteriviums zurück. Vor kurzem wurden Pollenkörner sogar im Valanginium gefunden (Brenner 1996). Die in den untersten Kreideschichten gefundenen Pollenkörner zeigen ziemlich von Anbeginn ein breites Spektrum an Ausprägungen. Einen Überblick über die Einteilung der Pollenkörner nach Art und Lage der Keimfalten und Keimporen gibt Abb. 1. Bis vor kurzem galt für die ältesten Pollenfunde folgende Situation (Abb. 2), wie sie auch in die Lehrbücher Eingang fand (vgl. Raven et al. 1988): Die Pollenkörner der ältesten Schichten hatten nur eine Keimfurche (sulcat), waren daher den Sporen der Farne und den Pollenkörnern der Gymnospermen ähnlich, und entsprachen dem Typ, wie er bei allen Monokotylen (einkeimblättrige Pflanzen) und den Magnoliopsida vorkommt. Pollenkörner mit drei Keimfurchen (tricolpat), wie sie bei der Mehrzahl der anderen Dikotylen bis heute typisch sind, treten vor "120 Millionen Jahren auf" (Raven 1988, S. 404; d. h. 7 Millionen Jahre später). Dies paßt gut in die verbreitete Vorstellung, daß die Magnoliopsida am Anfang der Angiospermen stehen, und es paßt ebenso gut zu der Beobachtung, daß auch bei den ältesten Makrofossilien die Magnoliopsida deutlich hervortreten. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Neueste Untersuchungen machen aber deutlich, daß die Lage weit komplizierter ist als oben dargestellt. So konnte Hughes (1994) mit Hilfe rasterelektronenmikroskopischer Aufnahmen zeigen, daß die Pollenstruktur sehr früher Funde weit vielgestaltiger ist als bis dahin angenommen. Außerdem ist die zeitliche Differenz zu den Schichten, in denen praktisch das gesamte Spektrum heutiger Formen zu finden ist, relativ unbedeutend. Vor allem mußten die herkömmlichen Vorstellungen durch die schon erwähnte Arbeit von Brenner (1996) revidiert werden. Anhand von fossilen Pollenkörnern aus verschiedenen Schichten der Unterkreide Israels konnte er erstmals zeigen, daß in den unteren Schichten Pollenkörner ohne Keimfurche (inaperturat) nicht selten waren, ja daß sogar die ältesten bisher bekannten Pollenkörner ausgerechnet diesem als abgeleitet geltenden Typ angehören, und daß dadurch der Anschluß an Gymnospermen und Farne problematisch wird. Welche Bedeutung dieser neuen Situation zukommt, kann man z.B. daran ersehen, daß die gerade erst vorgeschlagene deutsche Bezeichnung "Einfurchenpollen-Zweikeimblättrige" für die Magnoliopsida (Strasburger 1998) neu überdacht werden müßte, da die dortige Angabe, die Pollenkörner seien einfurchig oder gehörten zu davon abzuleitenden Typen, sehr fraglich geworden ist. Makrofossilien - AllgemeinesAngiospermenfossilien treten völlig unvermittelt in Schichten der Unteren Kreide auf (vor ca. 130 Millionen Jahren). Ihr Erscheinen ist geradezu explosionsartig. Die ältesten Blattreste stammen aus dem Barremium, Nachweise von Blüten und von Holz sind frühestens seit dem Aptium bekannt (Brenner 1996), der zweitjüngsten Serie der Unterkreide (s. Tab. 2). Ziemlich von Anfang an finden wir eine hohe Variabilität und eine hohe Spezialisierung bereits der ältesten Funde (vgl. Endress 1993, Crane et al. 1995). Soweit Blüten gut erhalten sind, gelingt es in der Regel, die Fossilfunde rezenten Familien zuzuordnen (Meyen 1987). Das heißt, ausgestorbene Familien, in denen am ehesten Zwischenformen zu erwarten wären, wurden nur selten gefunden. Der Vollständigkeit halber sei darauf hingewiesen, daß in älteren paläobotanischen Arbeiten die Zuordnung zu Gattungen oft fehlerhaft war, da sie überwiegend durch äußerlichen Vergleich mit den Blättern rezenter Formen erfolgte (Meyen 1987, S. 225). Moderne Untersuchungen, etwa aufgrund der Cuticularstrukturen, haben hier aber inzwischen Klärung gebracht. Die fossil überlieferte Gefäßpflanzenflora wird zu Beginn der Kreidezeit - was die Artenzahl angeht - etwa zu einem Drittel von Farnen und zu zwei Dritteln von verschiedenen Gruppen von Nacktsamern gestellt. Bedecktsamer sind artenzahlmäßig zunächst unbedeutend, machen aber bereits nach "nur" etwa 15 Millionen Jahren, zum Ende der Unterkreide (Albium), weit über die Hälfte der Gefäßpflanzenflora aus (Friis et al. 1987). Paläoökologische Studien (Abb. 3) zeigen, daß die Angiospermen in der Unteren Kreide bevorzugt an Ufern von Flüssen und Seen wuchsen, und daß sie im deutlichen Gegensatz zu ihrer taxonomischen Diversität, d.h. hohen Artenzahl, selbst noch in der Oberen Kreide nicht dominierten, sondern oft nur Unterwuchs darstellten (Taylor & Hickey 1996, Wing et al. 1993). |
|
|
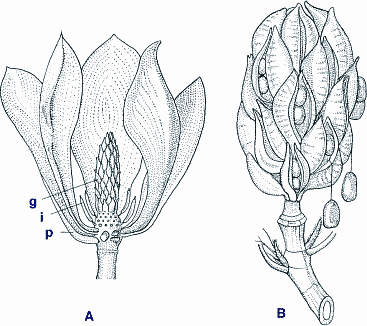
Die rasche Entfaltung der Angiospermen führte dazu, daß schon in der Unteren Kreide zahlreiche Pflanzenfamilien vertreten waren. Wegen der dichten zeitlichen Folge lassen sich daher aus den Fossilien keine Abstammungslinien ableiten. Es ist aber bemerkenswert, daß ein erheblicher Anteil der frühen Funde (einschließlich Pollen) zu den Magnoliopsida s.str. zu rechnen ist, die heute nur etwa 3% der Angiospermen ausmachen. Zu dieser Gruppe gehören Holzpflanzen wie etwa die bekannte Magnolie (Abb. 4), dem Lehrbuchbeispiel für eine primitive Angiosperme, ebenso wie krautige Pflanzen. Nach neueren kladistischen Untersuchungen sind die Magnoliopsida die Ausgangsgruppe aller Angiospermen (vgl. Tab. 1), von denen sich einerseits die heutigen Magnoliopsida, andererseits - wie oben erwähnt - die Monokotylen (Liliopsida) und die übrigen Dikotylen (Rosopsida) ableiten. Das Überwiegen der Magnoliopsida in den frühen Schichten ist insofern bemerkenswert, als diese Gruppe aus verschiedenen Gründen seit langem als ursprünglich diskutiert wird. |
|
|
Aus evolutionstheoretischer Sicht scheinen diese Verhältnisse sehr gut zusammenzupassen. Allerdings bleibt Erklärungsbedarf, wieso die ältesten Funde ausgerechnet nicht jene Merkmale zeigen, die man bei den Magnoliopsida für ursprünglich hielt. Für die Grundtypforschung stellt sich die Frage, wieso in den untersten Schichten andere Pflanzengruppen dominierten als heute. Es wäre zu überlegen, ob hier paläoökologische Aspekte zu berücksichtigen sind. Besonders beachtete Fossilfunde der letzten JahreIn den letzten Jahren gab es mehrfach Publikationen über die "ältesten Angiospermen", die vielfach auch in der Tagespresse aufgegriffen wurden. Einige der Funde sollen im Folgenden vorgestellt werden. Nicht eindeutig zu den Angiospermen zu rechnende Funde sind Irania hermaphroditica (vgl. Schweitzer 1989), Sanmiguelia aus der Trias mit unklarer systematischer Stellung (vgl. Taylor & Hickey 1996) und Pannaulika, ebenfalls aus der Trias (Cornet 1993; vgl. Taylor & Hickey 1996).
Chloranthacee (Taylor & Hickey 1990). Aptium, ca. 120 Millionen Jahre, Australien. Ältester Fund einer Angiosperme mit erhaltenen Blütenteilen. Pflanze krautig, Blätter wechselständig, in der Achsel jedes Blattes ein kleiner, komplex gebauter Blütenstand. Blüten sehr klein und ohne Blütenhülle. Die Fruchtblätter sind vermutlich miteinander verwachsen. Die Pflanze zeigt Merkmale, wie sie in verschiedenen rezenten Familien der Magnoliopsida vorkommen können. Zuordnung zu der auch rezent bekannten Familie der Chloranthaceae (Piperales, Magnoliopsida) möglich. Der Fund sorgte insofern für Aufregung, als er völlig den verbreiteten Vorstellungen einer primitiven Angiosperme widersprach und jener Hypothese Aufschwung gab, die den Ursprung der Angiospermen bei krautigen Vertretern mit unauffälligen Blüten sucht. Archaefructus liaoningensis (Sun et al. 1998, vgl. Crepet 1998). Oberer Jura, Yixianformation in China, ca. 140 Millionen Jahre (?). Dies wäre der älteste Angiospermenfund aufgrund von Blütenmerkmalen. Aber die Zugehörigkeit dieser Formation zum Jura ist nicht gesichert. Und auch die Zugehörigkeit zu den Angiospermen ist nicht eindeutig. Die Autoren stellen für Archaefructus eine eigene Unterklasse Archaemagnoliidae auf, da sie sich von allen anderen Angiospermen unterscheidet. Eine Blütenhülle fehlt. Die Zuordnung zu den Angiospermen erfolgt aufgrund der Fähigkeit der Pflanze, Samen aus zunächst völlig geschlossenen Früchten zu entlassen. Pollentragende Organe sind unbekannt. Blätter sind nur in wenig aussagekräftigen Resten vorhanden. Über den Habitus der Pflanze ist nichts bekannt. Crepet (1998) weist darauf hin, daß der Fund nicht zur Klärung der Frage der Verwandtschaft innerhalb der Angiospermen beiträgt, da die Kombination der Merkmale jegliche Verbindung zu irgendeiner bekannten Angiosperme unmöglich macht. Älteste Monokotyle (Gandolfo et al. 1998). Obere Kreide, ca. 90 Millionen Jahre. Winzige, millimetergroße, eingeschlechtige Blüten. Es wurden nur männliche Blüten gefunden. Aufgrund ihres Baues lassen sich die Funde der rezenten Familie der Triuridaceae zuordnen. Die Pflanzen sind chlorophyllfrei und auf die Ernährung durch Mykorrhiza-Pilze angewiesen (Myko-Heterotrophie). So ist dieser erste Fund einer einkeimblättrigen Pflanze ausgerechnet ein Beleg für eine hochspezialisierte Form. Das Fehlen von (fossil überlieferter) Angiospermen vor der KreideFossile Reste von Angiospermen fehlen völlig im Jura oder irgendeiner anderen tieferen Schicht. Jedenfalls erwiesen sich anderslautende Angaben stets als falsch oder mindestens sehr zweifelhaft (Crane et al. 1995). Diese Situation stellt sicher für jedes Modell über die Entstehung der Lebewesen eine Herausforderung dar und verlangt nach Erklärungen. Für die Evolutionstheorie ergibt sich die Frage, wo die zu erwartenden Zwischenformen existiert haben könnten. Denn von anderen in der Kreide nachgewiesenen Pflanzengruppen wird meist keine als Vorfahr der Angiospermen diskutiert (darauf wird im zweiten Teil in der nächsten Ausgabe eingegangen). Ein bekannter Versuch zur Lösung des Problems geht auf Axelrod (1952, 1961) zurück. Danach könnten Fossilien vor der Kreide deshalb fehlen, weil die frühe Evolution der Angiospermen außerhalb der Tiefebenen als normalen Fundorten von Fossilien stattfand. Entsprechend könnten die Angiospermen auf Anhöhen und in den Hochländern tropischer Regionen bereits einen großen Artenreichtum erlangt haben, ohne daß Pflanzenreste davon als Fossilien konserviert worden sind. Ein ähnliches Szenario wurde auch schon für die im Devon erstmals überlieferten Landpflanzen vorgeschlagen (vgl. Junker 1996). Ein anderer Lösungsansatz findet sich bei Schweitzer (1989, S. 163): "Die unmittelbaren Vorläufer der bedecktsamigen Pflanzen (der Angiospermen), die "Urangiospermen", müssen schon vor der Kreidezeit aufgetreten sein, da man bereits in der unteren Kreide ... viele Reste hochentwickelter Bedecktsamer findet. Es ist aber für einen Paläobotaniker außerordentlich schwierig, eine "Urangiosperme" als solche zu erkennen, weil die Übergänge zwischen nackt- und bedecktsamigen Pflanzen fließend gewesen sind, da sich alle Organe, auch die innerhalb einer Blüte, unabhängig voneinander entwickelt haben. So können Pflanzen mit hochentwickelten sterilen Organen primitive Blüten besessen haben und umgekehrt. Es wäre daher auch falsch anzunehmen, daß die Blüte einer "Urangiosperme" alle die Merkmale in sich vereinigt haben muß, die wir bei den Blüten der heutigen bedecktsamigen Pflanzen als primitiv ansehen. Vielmehr ist anzunehmen, daß es mehrere Urformen gegeben hat, die einander mehr oder weniger nahestanden und bei denen die einzelnen Organe verschieden weit entwickelt waren. Entscheidend ist nur, daß sie alle Merkmalskombinationen der bedecktsamigen Pflanzen in sich trugen." Solche Überlegungen haben in letzter Zeit eine gewisse Bestätigung durch kladistische Untersuchungen rezenter Formen gefunden, die die Entstehung der Angiospermen weit vor der Kreidezeit postulieren (siehe Folgeartikel in der nächsten Ausgabe). Das Fehlen der Angiospermen vor der Kreide verlangt auch im Grundtypmodell nach einer Erklärung. Wieso fehlt diese heute so dominante und vielgestaltige Gruppe im Jura, in der Trias, im Perm, Devon usw., obwohl es in diesen Schichten eine reiche Vegetation von Landpflanzen gab? |
Bevor wir uns näher mit der Frage einer möglichen Vorläufergruppe der Angiospermen beschäftigen, scheint es sinnvoll, uns einen Überblick über die Gymnospermen zu verschaffen (vgl. Tab. 1 und Abb. 6-8), da diese als einzige mögliche Ausgangsgruppe der Angiospermen diskutiert werden. |
|
Verglichen mit den Bedecktsamern (240 000 rezente Arten) sind die Nacktsamer (etwa 740 rezente Arten) vergleichsweise artenarm. Allerdings können die hierher zählenden Nadelbäume von der Individuenzahl her durchaus konkurrieren. Die Gymnospermen sind sehr heterogen. Seit einiger Zeit teilt man sie in zwei Unterabteilungen auf, die man für so unterschiedlich hält, daß sie innerhalb der Samenpflanzen gleichberechtigt neben die Angiospermen als dritter Unterabteilung gestellt werden (Tab. 1, Abb. 7). |
|
|
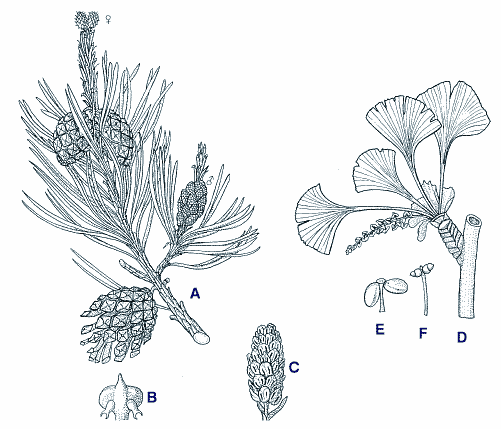
Allerdings gibt es auch andere Meinungen. So werten einige Autoren aufgrund von molekularbiologischen Daten der allerjüngsten Zeit die Gymnospermen trotz aller morphologischen Unterschiede als einheitliche, monophyletische Gruppe (vgl. Fröhlich & Parker 2000). 1. Coniferophytina = Nadel- und Gabelblättrige Nacktsamer (vgl. Abb. 7). Die weitaus bekanntesten und in Mitteleuropa auch die einzig vorkommenden Nacktsamer sind die Nadelbäume (Pinopsida). Hierhin gehören auch die früher z.T. abgetrennten Eibengewächse (20 Arten), und der zu einer eigenen Klasse (Ginkgoopsida) gehörige Ginkgobaum (mit nur einer Art) mit seinen bekannten Gabelblättern. Aus evolutionstheoretischen Überlegungen heraus pflegt man Nadelbäume als primitiv zu bezeichnen. Diese Sicht ist allerdings nur schwer mit ihrer hohen Vitalität und ihrer Konkurrenzfähigkeit in Einklang zu bringen. Fossil gibt es die Coniferophyten schon sehr lange. Die ausgestorbene Klasse der Cordaiten (Cordaitopsida) - hohe Bäume mit relativ großen, bandförmigen Blättern - hatte im Karbon und Perm eine ansehnliche Verbreitung, ist aber seit der Trias wieder völlig verschwunden. Die Pinopsida und Ginkgoopsida sind seit der Wende Karbon / Perm nachgewiesen und haben sich bis heute gehalten, wenn auch ein gewisser Teil der Arten ausgestorben ist. 2. Cycadophytina = Fiederblättrige Nacktsamer (Abb. 8). Die Blätter zeigen einen fiederigen Verlauf ihrer Leitbündel. Bei der mit 120 Arten artenreichsten Klasse der Cycadopsida (= Palmfarne) sind die Blätter selbst sogar gefiedert. Besonders interessieren uns die rezenten Gnetopsida (70 Arten) und die fossilen Bennettitopsida, da hier von vielen Autoren die Vorfahren der Angiospermen vermutet werden. Bei diesen beiden Klassen finden sich teilweise habituell und im Blütenbereich Anklänge an Angiospermen (vgl. Tab. 3). Zu den Gnetopsida gehören (neben seltenen ausgestorbenen Vertretern) drei sehr isolierte rezente Relikt-Gattungen: die merkwürdige Welwitschia (1 Art) aus Namibia mit nur einem Blattpaar und z.B. CAM-Stoffwechsel (als einziger Gymnosperme), die Gattung Gnetum (28 Arten) mit Laubblättern und u.U. Resten des zweiten Geschlechts in funktionell eingeschlechtigen Blüten, und schließlich die Gattung Ephedra (40 Arten), das sind Rutensträucher mit Schuppenblättern und Ansätzen zu Zwitterblüten und Insektenbestäubung. Bei den fossilen Bennettiteen gab es Arten mit zwittrigen Blüten und mit Tierbestäubung. |
|
|
Die Cycadeen und Bennettiteen waren im Jura und in der Kreide verbreitet. Während die Cycadeen sich bis heute gehalten haben, sind die Bennettiteen seit Ende der Unterkreide ausgestorben. Zu den Fiederblättrigen Nacktsamern werden auch die ausgestorbenen Samenfarne (Lyginopteridopsida) gezählt mit ihrem farnartigen Habitus und merkwürdigen Pollensack- und Samenanlagenträgern (vgl. Junker 2000). Sie hatten ihre Hauptverbreitung im Karbon und Perm, hielten sich aber noch bis zur Wende Jura / Kreide, d.h. sie sind noch vor der Angiospermenzeit ausgestorben. Obwohl sie den Angiospermen nicht besonders ähneln, werden sie von einigen Autoren als deren Ausgangsgruppe diskutiert, so auch in der gerade erschienenen Arbeit von Fröhlich & Parker (2000) aufgrund von DNA-Vergleichen an Regulationsgenen. |
Für phylogenetische Betrachtungen ist es wichtig zu wissen, wie klar zwei miteinander zu vergleichende Taxa voneinander unterschieden sind. Daher sei im Folgenden aufgezeigt, wie sich die Angiospermen von den Gymnospermen unterscheiden (Tab. 3). |
1. Die Samenanlagen stehen nicht frei, sondern sind in ein Fruchtblatt bzw. in einen Fruchtknoten eingeschlossen. 2. Im Zusammenhang damit wird eine Narbe ausgebildet. 3. Die Gametophyten (also die haploide Phase im Generationswechsel) sind auf wenige Zellen reduziert (Ausnahme: verschiedene Reduktionen bei Gnetopsida). 4. Außer der Eizelle wird noch eine weitere Zelle des Gametophyten befruchtet, die dann zur Ausbildung eines Nährgewebes (Endosperm) führt, das den Embryo ernährt. Hier ist aber darauf hinzuweisen, daß bei Ephedra und Gnetum in jüngster Zeit doppelte Befruchtung (wenn auch ohne Endospermbildung) nachgewiesen wurde (Friedman 1990, 1994). 5. Angiospermenblüten sind eingeschlechtig oder zwittrig, die der Gymnospermen fast immer eingeschlechtig und nur ausnahmsweise bei Ephedra und bei Vertretern der ausgestorbenen Bennettitopsida zwittrig. 6. Die Bestäubung erfolgt bei den Angiospermen durch Insekten oder Wind, bei den Gymnospermen (mit Ausnahme von Ephedra-Arten und von Bennettiteen) nur durch Wind. Man wertet die Windblütigkeit bei Angiospermen als abgeleitet. Strasburger (1998, S. 735) sieht einen Beweis dafür darin, daß viele windbestäubte Arten als Reste insektenbestäubter Arten noch z.B. Pollenkitt und Rudimente des jeweils anderen Geschlechts haben (Windbestäuber sind typischerweise eingeschlechtig). Man kann aber im Sinne des Grundtypmodells auch so argumentieren, daß viele Pflanzengruppen die Ausrüstung für beide Bestäubungsarten besitzen und entsprechend anpassungsfähig sind. Als Kuriosum sei folgender Satz aus Strasburger (1998) zitiert: "Gelegentlich kehren aber auch sekundär anemophile Sippen wieder zur Entomophilie zurück (z.B. Salix, Euphorbia)." Man vergleiche dazu den zweiten Teil dieses Beitrags in der nächsten Ausgabe. 7. In den Leitbündeln gibt es Siebröhrenglieder mit Geleitzellen, die aus einer Mutterzelle entstanden sind. 8. Angiospermen sind holzig oder krautig, Gymnospermen sind stets Holzpflanzen einschließlich Lianen. Im Holz fehlen den Gymnospermen die Tracheen, während die Angiospermen bis auf wenige Ausnahmen solche besitzen. Wie zu erkennen ist, gibt es einerseits eindeutige Unterschiede zwischen Angiospermen und Gymnospermen, andererseits sind einige bislang als angiospermentypisch gewertete Merkmale in letzter Zeit ansatzweise auch bei Gymnospermen festgestellt worden. Dies hat zum einen jenen Vorstellungen Auftrieb gegeben, die die Vorfahren bei den Gymnospermen suchen. Zum andern ist es aber bei konkreten Stammbaum-Rekonstruktionen nachteilig, wenn bisher angenommene Apomorphien (abgeleitete Merkmale) evtl. als Konvergenzen bzw. Symplesiomorphien gedeutet werden müssen (vgl. Taylor & Hickey 1996). In einem zweiten Teil - in der nächsten Ausgabe - sollen die unterschiedlichen Ansätze zur Lösung des Problems der Abstammumg der Angiospermen vorgestellt werden. Dabei werden auch neuere Ergebnisse molekularbiologischer Untersuchungen zur Sprache kommen. Schließlich soll im Detail diskutiert werden, welches nun konkret die Ausgangsgruppe der Angiospermen in evolutionstheoretischer Sicht sein könnte. | |||||||||||||||||||||||||||||||||
Literatur
|

|