|
|
|
|
|
|
|
Die pflanzlichen Überreste des Karbons gehören zu den am besten untersuchten pflanzlichen Fossilien. Ein Großteil der karbonischen Pflanzenfossilien gehört zu den Bärlappbäumen, den Lepidophyten (Ordnung Lepidodendrales). Sie wurden erstmals bereits in den zwanziger Jahren des 19. Jahrhunderts beschrieben. Bis zu 70% unserer Steinkohle besteht aus Resten dieser Pflanzengruppe (Kerp 1996), die großenteils im Laufe des Oberkarbons und schließlich vollständig im Perm ausgestorben ist. |
|
| Eine ungewöhnliche Konstruktion
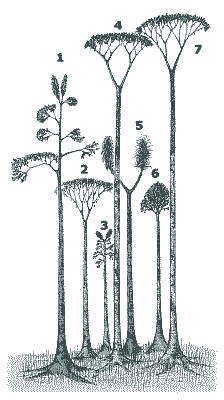
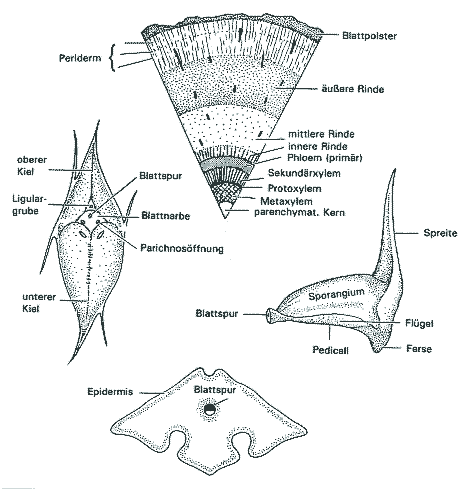
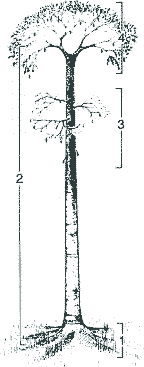
Was liegt näher als der Vergleich fossiler Lebewesen mit heute lebenden? Jeder Paläobotaniker muß zunächst von den Kenntnissen über heutige Formen ausgehen. Die Lepidophytenbäume - benannt nach den schuppenförmigen Blattpolstern, die die Rinde bedeckten (gr. lepis = Schuppe) - waren anatomisch, funktionell und ökologisch sehr verschieden von allen anderen Bäumen. Da sich die Paläobotaniker lange Zeit an heutigen Baumgestalten orientierten, wurde das Verständnis der Lepidophyten erschwert. Doch inzwischen ist klar, daß diese Pflanzengruppe so einzigartig gestaltet war, daß von einem speziellen "Bärlappbaum-Habitus" gesprochen werden muß (Phillips & DiMichele 1992, 560) oder auch von "Riesen-Kräutern" ("giant-herbs") die Rede ist (Bateman 1994, 543; vgl. Eggert 1961, 88). Die Lepidophyten demonstrieren, daß die meisten heutigen Pflanzen "bemerkenswert unrepräsentativ" für die ferne Vergangenheit sind (Bateman & DiMichele 1991, 195). Sie waren nicht "primitiv", sondern andersartig gestaltet. Morphologie der Luftstämme und -äste.Mit bis über 40 Meter Höhe (nach manchen Angaben bis 54 Meter) und bis zu 2 Meter Stammdurchmesser konnten die Lepidophyten es mit heutigen Bäumen (Gymnospermen und Angiospermen) ohne Weiteres aufnehmen. Gemessen an dieser Größe waren jedoch ihre Kronen meistens bemerkenswert schwach ausgebildet; im Extremfall (bei einigen Siegelbäumen) war die Krone gänzlich unverzweigt und bestand nur aus einem Blattschopf (war in diesem Fall also faktisch fehlend). Die Bäume waren stangenförmig gebaut (Abb. 1). In diesem Grundbauplan unterscheiden sich die baumförmigen Gattungen nicht grundsätzlich von "krautigen" (Phillips & DiMichele 1992, 564), die mindestens seit dem Unterdevon bekannt sind. Ungewöhnliche Stammanatomie.Nicht das Holz, sondern die Rinde und das Periderm sorgten für die Stabilität der Lepidophyten. Im Gegensatz zu den heutigen Holzpflanzen war die Wasserleitung nicht mit dem Stützgewebe gekoppelt. Der Rindenteil war viel stärker entwickelt als der Holzteil (Abb. 2 oben). Die Lepidophyten werden daher als "Rindenbäume" bezeichnet. Die Leitbahnen waren einfach ausgebildet (Proto- bzw. Aktinostelen; z. T. mit Mark). Sekundäres Phloem1 wurde erstaunlicherweise nicht gebildet (es wird w. u. noch deutlich werden, weshalb es nicht benötigt wurde). Vom Kambium (dem teilungsfähigen Gewebe) wurde also nur Xylem nach innen abgegeben, das höchstens wenige Zentimeter dick wurde. Jahresringe fehlten wie bei den meisten karbonischen Hölzern der nördlichen Halbkugel (Chaloner & Creber 1974). |
|
|
|
Die mächtige Rinde bestand gewöhnlich aus drei Regionen: einer schmalen Innenrinde aus kleinen Parenchymzellen, einer erheblich dickeren Mittelrinde mit großen, dünnwandigen Parenchymzellen und vielen Lakunen (Hohlräumen) sowie einer ebenfalls relativ breiten Außenrinde mit dickwandigeren und oft sklerenchymatischen (hartwandigen) Zellen. Bemerkenswert ist die Beobachtung, daß die Mittelrinde oft schlecht und in älteren Stämmen gar nicht erhalten ist. Ihre Zellen zerfielen vermutlich im Laufe des Lebens und hinterließen eine Höhlung (Taylor & Taylor 1993, 245, 257; vgl. Andrews et al. 1971, 9; Beck 1958, 452, 455). Blätter und Blattpolster.Die Blätter waren von typisch bärlappartigem Bau: nadelblattartige, schmale, meist kurze sog. Mikrophylle mit einer einzigen Blattspur. Die längeren von ihnen, die bis 1 m lang werden konnten, waren grasartig mit deutlich verbreiterter Basis (Stewart & Rothwell 1993, 135; der Begriff Mikrophyll ist also etwas unglücklich; er hängt mit phylogenetischen Hypothesen zusammen). Die Blätter fielen bald ab (bis auf die kleinen Blätter der Endzweige) und hinterließen Narben auf den charakteristischen arttypischen Blattpolstern, ohne daß allerdings ein echtes Trenngewebe ausgebildet wurde (Bateman et al. 1992, 514). Bei den Blattpolstern (Abb. 2 links) handelt es sich um Erhebungen auf der Stammoberfläche, die vermutlich dem Blattgrund der Mikrophylle entsprechen. Die Blattpolster waren langlebig, da sie mit dem Dickenwachstum Schritt hielten; sie dienten als Aufnahmeapparat für Regenwasser samt Flugstaub und Fäulnisprodukten; das Wasser wurde auf Ligulargruben hingeleitet (Zimmermann 1959, 185) Der Blattquerschnitt (Abb. 2 rechts) war meist rhombisch; auf der Unterseite befanden sich zwei Längsrillen mit Spaltöffnungen. Der Bau war also xeromorph, was auf den ersten Blick paradox zu sein scheint, wo doch die Pflanzen in feuchtem oder vermutlich sogar nassem Substrat (s. u.) wuchsen. Doch angesichts der Tatsache, daß das Leitsystem eher schwach ausgebildet war, ist dieser Befund letztlich doch verständlich. Zapfen und Verbreitung.
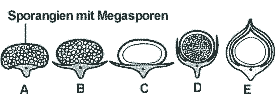
Die Zapfen befanden sich an den Enden von Laubsprossen und setzten sich aus spiralig (oder quirlig) stehenden Sporophyllen (sporangientragende Blätter) zusammen. Die Sporangien waren einzeln adadxial auf der Sporophylloberfläche angebracht (Abb. 2 unten). Die meisten Arten waren heterospor, wobei im Extremfall in den Megasporangien nur eine einzige funktionsfähige Megaspore gebildet wurde; bei manchen Gattungen wurde das Megasporangium mehr oder weniger von seitlichen Sporophyllflügeln umhüllt, so daß samenähnliche Gebilde ausgeprägt wurden (Abb. 3). Die Verbreitung erfolgte durch Heterosporen (Mega- und Mikrosporen), war also abhängig von Wasser. Manche Megasporen besaßen Ornamentierungen, die das Schwimmen ermöglichten oder erleichterten, z. B. hohle, unverzweigte Stacheln (Stublefield & Rothwell 1981, 632; Bateman et al. 1992, 540). Schwimmeinrichtungen sind auch bei Mikrosporen bekannt (Bateman et al. 1992, 540). Oft bildeten ganze Sporangien oder ganze Sporophylle die Verbreitungseinheit (Meyen 1987, 71); dabei waren die Sporophylle vielfach bootförmig ausgebildet und ideal zum Schwimmen geeignet (Leisman & Philipps 1979, 102; Gifford & Foster 1989, 153; Abb. 3). Begrenztes Wachstum.Trotz ihrer teilweise enormen Wuchshöhe entwickelten sich die Lepidodendren - wie bereits erwähnt - nicht in großkronige, langlebige Bäume, deren Kronen immer größer wurden. Nach einer gewissen Anzahl von Teilungen des Spitzenmeristems war dieses aufgebraucht und kein weiteres Wachstum mehr möglich. Die Bäume waren also nach einiger Zeit ausgewachsen und nicht mehr in der Lage, weiter zu wachsen und sich im Wuchs an eventuell sich verändernde Bedingungen anzupassen. Ihr Wachstum war demnach trotz baumförmigen Wuchses determiniert (Phillips & DiMichele 1992, 565f.). In dieser Hinsicht kann man diese Bäume mit Wirbeltieren vergleichen und von einem festen "Bauplan" sprechen (Bateman et al. 1992, 541). Es wird vermutet, daß die Bäume sehr schnell wachsen konnten, ziemlich kurzlebig waren, in vielen Fällen nur einmal (oder nur wenige Male) fruchteten und danach im Alter von nur 10 bis höchstens 15 Jahren abstarben. Photosynthese.Da die Blätter bald abfielen (s. o.), waren jeweils nur die äußersten Verzweigungen beblättert; nur die kürzeren Blätter an den Endverzweigungen fielen nicht mehr ab. Größere photosynthetische Bedeutung konnte den Blättern also nicht zukommen. Durch den frühen Laubfall und aufgrund der schütteren Kronen konnte das Licht jedoch weitgehend ungehindert auf den Stamm fallen, der von den bereits erwähnten Blattpolstern bedeckt war. Die Blattpolster besaßen Spaltöffnungen, waren also photosynthetisch aktiv. Dazu kommt die Luftversorgung durch sog. Parichnosbahnen. Diese Konstruktion ermöglichte (aufgrund des "light sharings" mit der Krone) der reifen Stammoberfläche, Photosynthese zu betreiben. Dadurch aber, daß überall an den Bäumen an Ort und Stelle Photosynthese erfolgen konnte, war eine Nährstoffleitung nicht erforderlich: nun wird verständlich, weshalb die Pflanzen kein sekundäres Phloem bildeten; sie benötigten es nicht.2 Die Verzweigung der Lepidophyten hatte denn auch nicht die Aufgabe, möglichst viele Anheftungsstellen für Blätter bereitzustellen, um dadurch eine große Oberfläche dem einfallenden Licht darzubieten, sondern sie diente möglicherweise fast ausschließlich oder mindestens überwiegend der Fortpflanzung, d. h. es handelte sich bei den Zweigen sozusagen um Zapfenanheftungsstellen (eventuell mit Ausnahme von Diaphorodendron scleroticum) (Phillips & DiMichele 1992, 578). Basale Organe.
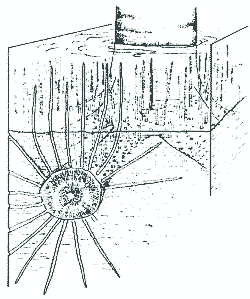
Sehr ungewöhnlich sind auch die basalen (unter"irdischen") Organe der Lepidophyten, die bis zu 15 Meter lang wurden (Taylor & Taylor 1993, 261). Bis heute behaupten sich verschiedene Auffassungen über die Natur dieser Organe (kurzer Überblick bei Frankenberg & Eggert 1969, 2f.). Es handelte sich jedenfalls nicht um Wurzeln, die anatomisch mit Wurzeln heutiger Pflanzen vergleichbar wären, sondern sie entsprechen deutlich den Luftsprossen. Außerdem fehlen Wurzelhaare und Wurzelhaube. Die Lepidophyten waren daher strenggenommen wurzellos. Die Päläobotaniker taten sich denn auch schwer, eine passende Bezeichnung für diese Pflanzenteile zu finden. Die größeren Äste werden wegen der mit Narben übersäten Oberfläche (s. u.) Stigmarien genannt und als Formgattung Stigmaria beschrieben. Der innere Bau (Abb. 4) gleicht histologisch und in der Zonierung der Rindengewebe dem Bau der Luftzweige, so daß die Stigmarien als umgebildete Äste betrachtet werden, die die Verankerung im Substrat bewirken (vgl. z. B. Eggert 1972, 93; eine Übersicht über den Bau bieten beispielsweise Frankenberg & Eggert 1969, 35-38).3 Der zentrale Markraum der Stigmarien war oft hohl. Wie bei den Luftsprossen war auch bei den Stigmarien die mittlere Rinde schlecht oder gar nicht erhalten; sie ist "sehr unvollständig bekannt", "die exakte Histologie ist nicht bekannt" (Frankenberg & Eggert 1969, 21, 35). Vermutlich war dieser Teil der älteren Stigmarien wie das Mark hohl. |
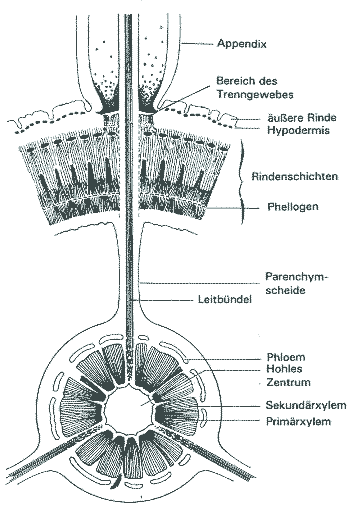 |
Abb. 4b: Stigmarie und Appendices. Querschnitt (nach Frankenberg & Eggert 1969) |
|
Die Stigmarien spitzen sich am Ende schnell zu und weisen eine Art konkaven Stöpsel aus Parenchym auf, der vielleicht die Funktion einer Schutzkappe hatte (Meyen 1987, 78; Rothwell 1984).
Die oft verwendete Bezeichnung "Wurzelträger" für die Stigmarien ist unglücklich, da sie keine Wurzeln tragen. Ihre Anhängsel, Appendices genannt, weisen im Bau und in der spiraligen Anordnung ebenfalls Beziehungen zu den Ästen von Luftzweigen auf. Sie wurden mit Hilfe eines Trenngewebes abgeworfen und hinterließen dann charakteristische Abbruchnarben (Stigmen), denen die Stigmarien ihren Namen verdanken (Abb. 5). Der Abwurf erfolgte auf erstaunlich ähnliche Weise wie der Blattfall heutiger Laubbäume (Frankenberg & Eggert 1969, 39). Die Innenrinde der Appendices zerfiel im Lebenzustand, so daß sie zu luftgefüllten Schläuchen wurden, die die (anfangs an einem parenchymatischen Septum aufgehängte) Stele umhüllten. Das Gefäßbündel in den Appendices ist bilateralsymmetrisch (Wurzeln haben sonst ein protostelisches radiärsymmetrisches Leitgewebe). 75-90% der basalen Biomasse bestanden aus den Appendices! Dies ist ein weiteres Argument dafür, daß die Appendices keine normalen Wurzelorgane waren (Phillips & DiMichele 1992, 569). Neuerdings wird sogar vermutet, daß das Stigmariensystem mit den Appendices photosynthetisch aktiv war. Durch das "light sharing" aufgrund der schwach ausgebildeten Kronen konnten oberflächennahe Appendices sehr wahrscheinlich genügend Licht einfangen, um es photosynthetisch zu nutzen. Sie konnten sich damit selbst versorgen (Phillips & DiMichele 1992, 567f.). Diese Vermutung wirft Fragen nach der Ökologie dieser Bäume auf, auf die weiter unten eingegangen wird.4 Alles in allem müssen die basalen Organe als Zweigsystem betrachtet werden, das für "Wurzelfunktion" modifiziert wurde (Frankenberg & Eggert 1969; vgl. Stubblefield & Rothwell 1981, 633). Ontogenese.Die Ontogenese der Lepidophyten verlief nach einem charakteristischen Muster: In einer frühen Phase, die als Epidogenese bezeichnet wird, erfuhr das primäre Gefäßsystem eine progressive Vergrößerung und Zunahme der Komplexität (Stewart & Rothwell 1993, 133). In der nachfolgenden Apoxogenese wurde mit jeder Dichotomie das terminale Meristem aufgebraucht, bis es schließlich zu klein war, um weitere Gabelungen zu ermöglichen, und verbraucht war (begrenztes Wachstum, s. o.). Ein vergleichbarer Vorgang ist von heutigen krautigen Lycopsiden bekannt; bei heutigen Gehölzen kommt ein solches Wachstumsmuster dagegen nicht vor. Als Folge davon konnten die kleineren Äste nicht in größere Zweige auswachsen (Taylor & Taylor 1993, 259). Die Blattpolster und die Blattspreite wurden zum Apex hin immer kleiner und weniger (Phillips & DiMichele 1992, 561). Die ersten ontogenetischen Stadien können mangels Fossildokumentation nur hypothetisch vermutet werden; es ist anzunehmen, daß sich zuerst das Stigmariensystem ausbildete. Da es wahrscheinlich photosynthetisch aktiv war, ist diese Annahme sinnvoll; außerdem würde es zunächst nahezu keinen Schattenwurf geben (Phillips & DiMichele 1992, 570f.). |
|
| Bauteile und Bauplanvariationen
Die Bärlappbäume bestanden aus bis zu vier Hauptbauteilen ("Module" nach Bateman et al. 1992, 543; Bateman & DiMichele 1991, 198): Stigmarien, Stamm, (verzweigte) Krone und Seitenzweige (Abb. 6). Je nach Vorhandensein und Kombination dieser Teile ergeben sich verschiedene Baupläne. Variation der Größe und der Menge an sekundärem Gewebe der vier Bauteile sowie Variation der Häufigkeit der Gabelungen der Seitenzweige und der Krone ermöglichte ein gewisses Spektrum von Wuchsformen (Bateman et al. 1992, 543; vgl. Abb. 1 und 7). Aufgrund des determinierten Wachstums gab es jedoch viel weniger Möglichkeiten im Vergleich zu den Angiospermen, aufgrund von Umwelteinflüssen die Wachstumsarchitektur anzupassen (Bateman et al. 1992, 543; Bateman & DiMichele 1991, 198). Einige Gattungen besaßen einen scheinkrautigen Wuchs ("pseudoherbs"), eine Wuchsform, die heute in dieser Form nicht vorkommt (Abb. 7). Wesentliche Merkmale sind die Beschränkung von Holz und Periderm auf die Hauptachse und die Stigmarien, ein sehr kurzer Stamm, der die Zweige nicht deutlich über den Erdboden erheben kann (Bateman & DiMichele 1991, 199), und radiärsymmetrische Zweige ohne Adventivwurzeln (vgl. Bateman & DiMichele 1991, 200). Echte prostrat wachsende Kräuter besitzen dagegen eine bilateralsymmetrische Achse und Adventivwurzeln.
|
|
| Ökologische BetrachtungenWichtige Merkmale der Lepidophyten sollen nun unter ökologischen Gesichtspunkten aufgegriffen und kommentiert werden. Einige Eigenschaften sind typisch für Pionierpflanzen:
Diese Eigenschaften sind auffällig vergleichbar mit den Lebensstrategien einjähriger Angiospermen-Kräuter. Aber hier handelt es sich um Bäume oder niederliegende "Gehölze" ("pseudoherbs"). Von Pionierpflanzen sollte man weiterhin erwarten, daß sie sich effektiv vegetativ fortpflanzen können, doch das ist bei den Lepidophyten nicht der Fall; vegetative Vermehrung ist bei ihnen unbekannt (Philipps 1979, 240). Weitere bemerkenswerte Eigenschaften der Lepidophyten deuten (zusätzlich) in eine andere Richtung. Dem aufmerksamen Leser dürfte nicht entgangen sein, daß recht viel von Anpassung ans Wasser bzw. von Eigenschaften, die viel besser (oder nur) zu Wasserpflanzen passen als zu Landpflanzen, die Rede war. Im Einzelnen:
Neben diesen deutlichen Wasser-Lebensstrategien sind weitere Merkmale am ehesten bei einer Lebensweise im Wasser verständlich:
Insgesamt drängt sich ein Bild auf, das gewöhnlich nicht diskutiert wird, für das aber neben den genannten anatomischen Merkmalen auch geologische Indizien angeführt werden können (vgl. Scheven 1986): die Lepidophyten befanden sich im Wasser, ohne festen Kontakt zu einem Boden zu haben. Sie waren nicht nur großenteils im Wasser, sondern ganz im Wasser: schwimmende Wälder. Insbesondere der Bau der Stigmarien und der Appendices läßt kaum eine andere Deutung zu.5 Diese Vorstellung ist nicht neu. Sie wurde am Ausgang des letzten Jahrhunderts besonders von Kuntze vertreten (vgl. Kuntze 1884, 1895; s. Kasten) und in modifizierter Form von Scheven (1986) neu in die Diskussion gebracht. Das Schwimmwaldkonzept fand jedoch wenig Beachtung (vgl. die Kritik von Gastaldo 1985). In einem Folgebeitrag sollen evolutionstheoretische Studien vorgestellt und einer kritischen Beurteilung unterzogen werden. |

|