 |
|
von Reinhard Junker
Studium Integrale Journal
23. Jahrgang / Heft 2 - Oktober 2016
Seite 75 - 82
|
 |
Zusammenfassung: Im Rahmen evolutionsbiologischer Ansätze wird die Entstehung von Vogelfedern und Vogelflug nach wie vor kontrovers diskutiert. Den Grund dafür bringen Proctor & Lynch (1993, 88) so auf den Punkt: „Federn sind ein bisschen zu perfekt – das ist das Problem.“ Diese Perfektion zeigt sich nicht nur in den Einzelteilen der Vogelfeder, sondern in den komplexen Wechselbeziehungen der Teile des gesamten Flugapparats – für evolutionäre Hypothesen eine harte Nuss.
|
|
|
Einleitung
Die Entstehung der Vogelfeder und des Vogelflugs gehört zu den prominentesten Themen der Ursprungsforschung in der Biologie (Prum & Brush 2003). Im Rahmen evolutionsbiologischer Ansätze ist die Entstehungsweise bis heute umstritten. Die Mehrheit der Forscher sieht die hypothetische evolutionäre Entstehung der Feder nicht im Kontext der Entstehung des Fluges. Vielmehr sollen Federn ursprünglich andere Aufgaben gehabt haben (Wärmedämmung, Balz u. a.) und erst später zusätzlich für den Flug verwendet worden sein. Nur wenige Forscher sind der Auffassung, dass Federn von Beginn an für den Flug evolviert seien.
Sowohl neuere Befunde aus der Embryologie und Entwicklungsgenetik als auch zahlreiche neue Funde von Dinosauriern mit faserigen oder fiederigen Körperbedeckungen haben in den letzten etwa 20 Jahren die Datenbasis für Modellierungen der Entstehung von Vogelfedern und Vogelflug erheblich erweitert. Theoretische Szenarien zur Entstehung von Federn und Flug können daher einem sehr viel umfangreicheren und detaillierteren „Faktencheck“ unterworfen werden. Ob damit auch mehr Klarheit über die Entstehung dieser hervorstechendsten Merkmale der Vögel erreicht wurde, soll in diesem Artikel und nachfolgenden Beiträgen untersucht werden. In vielen Fachartikeln und noch mehr in populären Darstellungen wird einerseits häufig der Eindruck erweckt, als sei mit den zahlreichen Funden „befiederter“ Dinosaurier der evolutive Weg zur Vogelfeder und zum Vogelflug in Grundzügen geklärt. Andererseits hat in den letzten Jahren auch die Anzahl der unterschiedlichen Modelle zur Entstehung des Vogelflugs zugenommen – ein ziemlich sicheres Symptom dafür, dass diese Frage nicht beantwortet ist.
Wenn aufgeklärt werden soll, auf welchem Weg Vogelfeder und Vogelflug entstanden sein könnten, dann müssen das Erklärungsziel und die Anforderungen an eine gültige Erklärung klar beschrieben sein. Das heißt: Grundlegende Voraussetzungen hinsichtlich der strukturellen Anforderungen an Federn, einschließlich ihrer Verankerung im Körper, ihrer Beweglichkeit und Steuerung, ihrer Ontogenese, aber auch an das gesamte Federkleid und die Anatomie des Tieres als notwendige Bedingungen für die Flugtauglichkeit von Federn bzw. die Flugfähigkeit des Vogels allgemein müssen definiert und beachtet werden. Im Folgenden werden dazu einige Punkte detailliert betrachtet.
|
Vogelfedern sind die komplexesten Integumentstrukturen (Körperbedeckung) des Tierreichs und erfüllen vielfältige Funktionen. In diesem Beitrag steht ihre Funktion als Teil des Flugapparats im Fokus. Bei aller Perfektion ermöglichen Federn alleine jedoch bei weitem nicht die Flugfähigkeit. Neben geeignetem Federmaterial und detailliertem Feinbau ist auch eine passende Verankerung der Federn in der Haut unabdingbar, ebenso ein komplexes Geflecht von Federmuskulatur und Sinnesorganen für die Bewegung der Federn. Außerdem muss insgesamt ein funktionsfähiges Federkleid ausgebildet sein, vielfältige Steuerungsmechanismen und Koordination der Bewegungen sind erforderlich, eine entsprechende Gehirnorganisation und anderes mehr.
Entscheidend für die Funktionsfähigkeit von Federn sind auch Struktur und Komplexität des Follikels, insbesondere in der ontogenetischen Entwicklung und bei der Mauser. Da Federn durch verschiedene äußere Einflüsse abgenutzt werden, muss ihr Austausch in der Mauser geregelt sein – ebenfalls ein komplizierter Vorgang. Schließlich muss eine beständige Gefiederpflege mit den dafür erforderlichen Pflegemitteln und Verhaltensweisen garantiert sein.
Aus diesen Gründen weisen viele Forscher auf den Aspekt der Synorganisation bei Federn hin. Die einzelnen Module und Ebenen (Baumaterial bis Verhalten) können nicht isoliert voneinander verstanden werden und auch nicht isoliert entstanden sein. In Summe haben wir mit dem Federkleid eine Gesamtorganisation vor uns, die bezüglich der Flugfähigkeit nichtreduzierbar komplex erscheint und ein klares Design-Indiz darstellt.
|
|
|
Aufbau, Material, Pflege
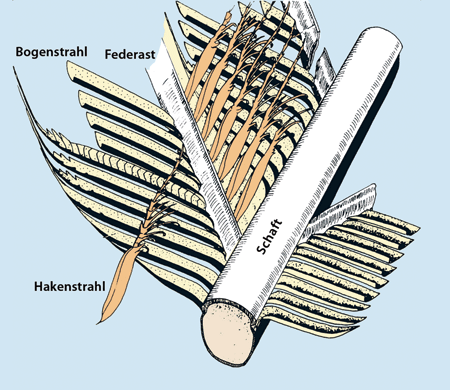 |
| Abb. 1: Schaft, Federäste, Haken- und Bogenstrahlen. (Aus Junker & Scherer 2013) |
|
Federn erfüllen gleichzeitig mehrere Funktionen, vor allem ermöglichen sie im Zusammenwirken mit vielen anderen morphologischen und verhaltensbiologischen Voraussetzungen die Flugfähigkeit, dienen aber auch der Thermoregulation, dem Tastsinn, der Brutpflege, dem physikalischen Schutz und der Wasserabstoßung und besitzen Signalfunktion (z. B. hinsichtlich Balz, Territorialverhalten). „Mehr als 20 Funktionen werden von Federn maßgeblich oder teilweise bestritten“ (Kämpfe 2003, 42). Dafür müssen der Aufbau und das Material besondere Anforderungen erfüllen. Federn sind sehr filigrane Gebilde und trotz ihrer Leichtigkeit höchst robust und flexibel (Prum & Brush 2003, 34). Der Leichtbau ist unverzichtbar für die Flugfähigkeit. Der Federschaft ist dementsprechend im ausgewachsenen Zustand hohl und die Federproteine (beta-Keratine, eine bedeutende Klasse der Strukturproteine) ermöglichen ein geringes Gewicht. Die Federkeratine sind faserig und bilden untereinander feste Strukturen (Prum & Brush 2003).
Jede Feder besteht aus über 1 Million Haken- und Bogenstrahlen (Bergman 2003, 34; vgl. Abb. 1-3). Ohne Haken an den Hakenstrahlen (wie zum Beispiel beim Vogel Strauß) sind die Federn nicht flugtauglich. Die Verhakung der Hakenstrahlen mit den gegenüberliegenden Bogenstrahlen kann bei Bedarf (z. B. bei mechanischer Beanspruchung) gelöst und in einer Art Reißverschlussverfahren wieder verknüpft werden. Aufgrund des Verhakungsmechanismus verhalten sich Schwungfedern trotz feiner Untergliederung der Federfahne wie eine geschlossene Tragfläche und sind erstaunlicherweise kaum luftdurchlässig. Eventuelle Risse können aufgrund dieses Bauprinzips mit Hilfe des Schnabels schnell repariert und die Geschlossenheit der Fläche wiederhergestellt werden.1 „Nur durch diese Eigenschaft ist eine Feder als Element eines effizienten Flugapparats überhaupt tragbar“ (Peters 1994, 404). „Gegenüber einer tatsächlichen Festfläche hat die Technologie einer Federfahne einen beträchtlichen Vorteil. Sie besitzt sozusagen vorgefertigte Trennlinien, an denen sie bei Extrembelastungen oder beim Anstoßen an irgendwelche Hindernisse aufreißen kann. Durch diesen Effekt des ‚eingebauten Nachgebens‘ wird verhindert, daß sich die Feder bei jeder akuten Überbelastung selbst zerstört“ (Nachtigall 1985, 38).
 |
| Abb. 2: Aufbau einer Konturfeder. (Nach Proctor & Lynch 1993 bzw. Lucas & Stettenheim 1972) |
|
Das Bogen-Hakenstrahlen-System als strukturelle Voraussetzung würde allerdings noch nicht eine dauerhafte Flugtauglichkeit der Feder garantieren. Es sind auch Instandhaltung, Wartung und Pflege durch eine Art Schmieröl erforderlich, damit die Gelenke an den Bogen- und Hakenstrahlen intakt bleiben, Unebenheiten geglättet werden und somit die Funktionalität der Federn erhalten bleibt. Das Gefiederfett entnehmen die Vögel aus der im Bereich des Steißes befindlichen Bürzeldrüse. Mit Hilfe des Schnabels verteilen die Vögel das Fett, was gleichzeitig der Imprägnierung dient und die Federn geschmeidig erhält. Für die Gefiederpflege benötigen Vögel viel Zeit2, denn Federn müssen ständig einzeln von Schmutz und Parasiten gereinigt werden, damit sie in gutem Zustand bleiben und gegen Bakterien und Pilze geschützt sind.3
 |
| Bei flugunfähig gewordenen Vögeln gehen die Asymmetrie und der Verhakungsmechanismus der Federn schnell verloren. |
 |
|
 |
| Abb. 3: Details eines Federastes. (Nach Proctor & Lynch 1993 bzw. Lucas & Stettenheim 1972) |
|
Mikrostruktur. Lingham-Soliar & Murugan (2013) und Lingham-Soliar (2014) haben die Mikrostruktur von Federn detailliert untersucht und ein komplexes, verschachteltes System von überkreuzten Keratinfasern mit einem Durchmesser von 100-800 nm in der Proteinmatrix im Federschaft und in den Federästen entdeckt. Die Fasern sind in verschiedenen Lagen des Epicortex (seitliche Wände des Schafts) gegenständig angeordnet, und sie bilden eine ununterbrochene strukturelle Verbindung zwischen dem Cortex der Rachis und der Federstrahlen. Die seitlichen Wände, von denen die Federstrahlen (Bogen- und Hakenstrahlen) ausgehen, sind sehr viel dünner als die Ober- und Unterseite. Dieses ausgeklügelte System im Bau und in der Anordnung der Keratinfasern ermöglicht die paradoxen, aber unabdingbaren mechanischen Qualitäten von Federn: Starrheit und Festigkeit einerseits und Leichtheit, Biegsamkeit und Elastizität andererseits.4 Die Fasern des beta-Keratin kombinieren Bruchsicherheit, Steifheit, Dehnbarkeit und Stauchbarkeit, Drehbarkeit und Reißfestigkeit (Lingham-Soliar 2014, 329).
Federform. Eine weitere notwendige Voraussetzung für die Flugtauglichkeit von Federn ist ihre äußere Form (Umriss). Um Auftrieb zu ermöglichen, muss die Luft oberhalb der Oberfläche der Feder schneller strömen als unterhalb (Bernoulli-Effekt). Dies wird dadurch erreicht, dass die Außenfahne, die gegen die Luftströmung steht, schmaler und steifer ist als die Innenfahne.5 Nur asymmetrische Federn sind flugtauglich. Bei flugunfähig gewordenen Vögeln gehen die Asymmetrie und der Verhakungsmechanismus der Federn schnell verloren.
|
|
Beweglichkeit und Steuerung
 |
Zur Funktionalität der Federn gehört auch ihre Beweglichkeit. Sie wird dadurch ermöglicht, dass Federn in die Körperoberfläche eingesenkt sind (Abb. 4, 8). An einer eingesenkten Tasche, dem Federfollikel, setzt ein komplexes Geflecht von Muskeln und elastischen Sehnen an (Abb. 5, 6). Dabei sind mehrere benachbarte Follikel (und damit die Federspulen, Calami, die eingesenkten Teile der Federn) miteinander verbunden (Bock 2000a, 479).6 Kämpfe (2003, 41f.) beschreibt diese Muskeln wie folgt: „Zur Beweglichkeit der Federn heften sich am Federfollikel glatte Muskeln an und verbinden benachbarte Follikel. Die Faserzüge kreuzen sich und bilden Netzwerke. Hinzu treten quergestreifte Hautmuskeln. Dabei ist interessant, dass die Hebermuskeln der Feder weniger kräftig sind als die Depressormuskeln. Letztere werden nicht nur als Antagonisten der Heber, sondern vor allem zum Stabilisieren bei Luftströmungen benötigt. Ein gut entwickelter Fettkörper in den Vertiefungen der Körperoberfläche wirkt bei der Bewegung der Federn wie ein hydraulisches Kissen und trägt zur optimalen Federbewegung und -stellung bei.“ Durch dieses Geflecht von kräftigen Muskelsträhnen können die Einzelfedern aufgerichtet, gedreht und wieder niedergezogen werden (Burckhardt et al. 1979, 13). Bei Konturfedern (nicht bei Daunenfedern) ist somit ein komplexes hydraulisches skeleto-muskuläres System mit Fettkörper ausgebildet (Homberger & De Silva 2000, 560). Peters (2001, 397) beschreibt dieses System so: „Die Follikel der Konturfedern heutiger Vögel sind durch ein sinnvolles System glatter Muskeln verbunden. Die Bewegungen der Federn ergeben sich aus einem komplexen Zusammenwirken dieser Muskeln mit einem Hydroskelett aus elastischen Faszien und inkompressiblen Fettpolstern.“ Jeder Federfollikel ist „von 2-6 solcher glatten Muskeln gehalten, wobei jeder Muskel jeweils in mehreren Bündeln zwischen 2 Federfollikeln liegt und mit diesen Follikeln verbunden ist“ (Burckhardt et al. 1979, 13). Lediglich den Fadenfedern fehlt die Muskulatur (Maderson et al. 2000, 705).7 Deren Rezeptoren informieren den Vogel über die Stellung der Federn (Bossert 2000). „Die Federn sitzen normalerweise sehr fest in ihren Follikeln, sowohl in der Wachstumsphase als auch nachher: Die Federfollikel werden durch die Federmuskeln und ihre elastischen Sehnen zusammengezogen ...“ (Burckhardt et al. 1979, 13).
Muskeln benötigen Innervierung und Energiezufuhr über das Blut; die Steuerung der Muskeln bedarf auch der Information über die Stellung der Federn; dafür sorgen besondere Tastsinnesorgane: „Zu seiner Steuerung ist die nervliche Ausstattung des befiederten Integumentes optimal: An den Muskeln und Federbälgen sitzen zahlreiche Sensoren: Tastkörperchen sind besonders zahlreich. Sie registrieren vor allem die Bewegungen der Konturfedern (Abb. 7). Die Erregungsmuster werden in Rückenmark und Kleinhirn verarbeitet“ (Kämpfe 2003, 42). Homberger & de Silva (2000, 564) sprechen von einem Feedbackmechanismus durch ein kompliziertes Nerven-Netzwerk mit Tastrezeptoren und Innervation für die Bewegung.8
Abb. 4: Schema einer fortgeschrittenen Federanlage, teilweise aufgeschnitten (nach Peters 2001). Der Follikel entsteht durch Einsenkung einer Papille in die Haut. Dabei vermehren sich Zellen in einem Ring um die Federanlage herum. Vom Epidermalkragen werden Zellen nach oben abgegeben, die zu Teilen der Feder werden. Die Feder entwickelt sich zunächst als Zylinder in der anfänglich noch geschlossenen Federscheide. Der Epidermalkragen teilt sich beim Hochwachsen gleich in Säulen, aus denen später die Federäste werden. Die Federentwicklung verläuft fast komplett anders als die Entwicklung von Reptilschuppen. (Aus Junker & Scherer 2013) |
|
|
|
Federkleid
 |
| Abb. 5: Quervernetzte Muskulatur setzt an den Follikeln an. (Nach Peters 2001; in Anlehnung an Osborne 1968) |
|
Flugtaugliche Federn mit den beschriebenen Bewegungs- und Steuermöglichkeiten würden aber noch keine Flugfähigkeit ermöglichen. Die Federn müssen auch sinnvoll auf der Körperoberfläche verteilt sein. Dementsprechend sind Federn nicht wahllos auf der Körperoberfläche angeordnet.
Befiederte Bezirke der Körperoberfläche werden als Federfluren bezeichnet, die unbefiederten als Federraine (Abb. 9); die Federn stehen relativ weit auseinander (Martin 2008, 469). Neben den an den Follikeln ansetzenden Muskeln sind auch Muskelbezirke aus willkürlich beherrschten Hautmuskeln zur Bewegung der Federfluren notwendig. Weitere Hautmuskeln dienen als Spanner der Flughäute, als Spreizmuskeln der Flügelschwingen und als Aufrichter der Schwanzfedern (Burckhardt et al. 1979).10
|
|
Ontogenese
Das Bild kann online nicht zur Verfügung gestellt werden. |
| Abb. 6: Muskel- und Fasergeflecht im Bereich der Follikel. (Nach Burckhardt et al. 1979) |
|
Federn wachsen ganz anders als Reptilschuppen. Schuppen bilden sich aus Hautfalten, Federn dagegen erheben sich aus einer röhrenförmigen Hauteinsenkung (Prum & Brush 2003, 35). Nur die ersten Stadien der Schuppen- bzw. Federontogenese – die Bildung einer Papille (Aufwölbung der Haut; Abb. 8) – sind vergleichbar. Die Federanlagen bilden sich bereits etwa ab dem 5. Lebenstag im Ei.
|
| „Federwachstum ähnelt mehr der Art, wie ein Bildhauer eine Statue aus einem Marmorblock herausmeißelt.“ |
|
|
Die Papille wächst zu einem Zapfen aus, um den sich in der Folge ein tiefes Ringfeld bildet, wodurch der Zapfen eingesenkt wird – der Follikel (bzw. Federbalg) entsteht. In der weiteren Folge stellt sich der Zapfen schräg. Die Epidermis im Zapfen differenziert sich in die Federscheide (äußere Malpighische Schicht, sie fungiert als verhornende Schutzschicht) und in die Federanlage (innere dreiteilige Schicht). Der innere Teil des Zapfens – die Pulpa (das Mark) aus Hautgewebe – versorgt die wachsende Feder mit Nährstoffen und wird später hohl.
Nun bilden sich die Rachis und die Federfahne sowie ggf. die Federstrahlen in einem komplizierten Prozess. „Die Wachstumsmuster folgen … einer besonders komplizierten Choreografie“ (Prum & Brush 2003, 35); „eine komplexe Abfolge von Ereignissen vermittelt der Feder ihren Charakter als entweder weiche Daunenfeder oder als Flugfeder mit einer steifen Fahne“ (Proctor & Lynch 1993, 86). Die Federn wachsen von der Basis her; im dort befindlichen Epidermalkragen (Bildungszone) teilen sich Keratinzellen und schieben ältere Zellen nach oben, die zu Teilen der Feder werden. Die am weitesten differenzierten Teile der wachsenden Feder befinden sich also oben. Die Feder entwickelt sich zunächst als Zylinder in der anfänglich noch geschlossenen Federscheide. Der Epidermalkragen teilt sich beim Hochwachsen gleich in Säulen (innerer Ring), aus denen später die Federäste werden. Bei den Konturfedern entsteht der Schaft dadurch, dass „die unteren Säulenenden an einer Seite der Röhre eine nach der anderen miteinander verschmelzen. Gleichzeitig schieben sich an einer Stelle der Gegenseite von unten neue Säulen nach. Sie drücken dabei die schon vorhandenen Säulen schräg zur Seite, sodass sich diese spiralig um die Röhre legen“ (Prum & Brush 2003, 36). Schließlich reißt die schützende Federscheide oben auf und entlässt die Feder.
Steve Hunter beschreibt den Vorgang der Federbildung durch ein interessantes Bild: „Federwachstum ähnelt mehr der Art, wie ein Bildhauer eine Statue aus einem Marmorblock herausmeißelt – die Statue von David war schon im Felsen, Michelangelo entfernte lediglich alles, was nicht David war. In ähnlicher Weise startet eine Feder nicht als Schaft, aus dem Zweige sprießen, sie beginnt vielmehr als sich verjüngender Zylinder, und Zelltod formt die Strukturen.“11
Lin et al. (2006) unterscheiden bei der ontogenetischen Federbildung fünf Prozesse, in denen allesamt komplexe molekulare Vorgänge beteiligt sind: Bildung der Makrostrukturen, Bildung der Feinstrukturen, Formbildung innerhalb der Einbuchtung, Formbildung des Folikels, Regeneration.12
 |
| Abb. 8: Vier ontogenetische Stadien der Federentwicklung. Näheres im Text. (Nach Proctor & Lynch 1993) |
|
|
|
Mauser
Federn nutzen sich im Laufe eines Jahres ab und müssen daher regelmäßig erneuert werden; diesen Vorgang nennt man Mauser. Die alten Federn fallen aus den Follikeln heraus, und es wachsen aus den gleichen Follikeln neue Federn nach. Die Bildungszone wird dabei wieder aktiv. Die Mauser erfordert einen periodischen Wachstumsstillstand und eine Desorganisation des Epidermalkragens (Wachstumszone, vgl. Abb. 8), anschließend die Reorganisation des Kragens und Wiederaufnahme des Wachstums der neuen Feder (Prum 1999, 301).13 Es ist offensichtlich, dass dieser Vorgang hohe Programmierkunst erfordert. Wann die Mauser erstmals entstand, ist unbekannt (Prum 1999, 301).14
Nicht nur der Ersatz der einzelnen Federn erfolgt koordiniert, sondern auch die Abfolge des Austausches der Federn: „Wenn sich Vögel mausern, ersetzen sie ihre Flugfedern gewöhnlich nach einem festgelegten Schema, das von Art zu Art variiert. Die meisten wechseln ihre Arm- und Handschwingen beiderseits in Paaren, um während der Mauser flugfähig zu bleiben. Viele Wasservögel jedoch mausern alle Handschwingen gleichzeitig und sind dann für einige Wochen flugunfähig“ (Kamphuis 2008, 29).
|
|
Federn als neuartige Gebilde
Der Vergleich der Ontogenesen von Reptilschuppen und Vogelfedern lässt heute kaum einen Zweifel daran zu, dass Federn neuartige Gebilde sind – im Rahmen des Evolutionsparadigmas wird von einer evolutionären Neuheit gesprochen. Genauer gesagt kommen mehrere Neuheiten zusammen: der Follikel, die röhrenförmige Federanlage und die komplexe Federstruktur (Prum & Brush 2002, 265; vgl. Stettenheim 2000, 471).15 Dabei hat der Follikel besondere Bedeutung. Er ist ein komplexes Organ, dessen Aufbau die Bildung und das Wachstum der Federn ermöglicht. Die Positionierung des Follikels und die Regulation der Entwicklung innerhalb des Follikels werden durch eine komplexe Abfolge von Induktionen und Kommunikation zwischen Haut und Epidermis festgelegt. Mit wenigen Ausnahmen werden alle Follikel, die die Federn im Laufe des Lebens eines Vogels hervorbringen, während der ersten 12 Lebenstage noch im Ei angelegt (Prum 1999, 293).16 Dabei entwickeln sich die verschiedensten Federtypen aus derselben Grundstruktur des Follikels; dieses Organell kann somit sehr vielseitige Strukturen hervorbringen (Prum 1999, 301).17 Obwohl also der Aufbau der Follikel (abgesehen von ihrer Größe) sehr einheitlich ist, kann daraus eine enorme Vielfalt von Federtypen hervorgehen.
|
|
Synorganisation
Führende Wissenschaftler weisen darauf hin, dass Federn nicht als isolierte Strukturen betrachtet werden dürfen, sondern als Teil eines komplexen Organs, des Integuments (Bock 2000, 479).18 Überlegungen zur Federevolution müssen daher auch die Verankerung der Federn in der Körperoberfläche und ihre Steuerung berücksichtigen (Homberger & De Silva 2000, 554).19 Die Körperhülle besteht in federtragenden Bezirken aus drei Schichten: einem hydraulischen skeleto-muskulären Apparat der Federn, einem System von Muskeln in der Unterhaut und einem hydraulischen System aus Fettgewebe (Homberger & De Silva 2000, 556).20 Die Haut- und Unterhautmuskeln bilden ein integriertes Muskelsystem, in welchem jeder Muskel eine spezifische Rolle spielt, um die korrekte Position und Anordnung der Federfluren auf der Oberfläche des sich bewegenden Körpers sicherzustellen, was notwendig für die einwandfreie Funktionsweise des hydraulischen skeleto-muskulären Apparats der Federn ist (Homberger & De Silva 2000, 560).21
|
| Es ist nicht realistisch, eine Evolution der Federn unabhängig vom Vogelintegument und aller seiner quervernetzen Strukturen zu diskutieren. |
|
|
Den Aspekt der Synorganisation stellt auch Bock (2000, 479) heraus: „Federn existieren nicht als isolierte Strukturen auf der Oberfläche des Vogelkörpers, sondern sind zusammen mit anderen Strukturen miteinander verbunden, um das Federkleid zu bilden. Das Gefieder bildet zusammen mit der Haut, einschließlich einer Fettschicht, Haut- und Unterhautmuskeln, Bändern und Sinnesorganen … eine vernetzte Folge verschiedener Merkmale – ein Organ.“22
 |
| Abb. 9: Federfluren: Befiederte Bereiche der Haut. (Wikimedia Commons, CC BY-SA 3.0) Burckhardt et al. 1979) |
|
Aus diesen Gründen ist es nicht realistisch, eine Evolution der Federn unabhängig von der Evolution des Vogelinteguments und aller seiner quervernetzen Strukturen zu diskutieren (Bock 2000, 479).23 Die Evolution der Vogelfedern sei von größeren damit verbundenen Innovationen ihrer Körperverankerung, Färbung, Anordnung, Betätigung, Wachstum und Mauser begleitet, so Stettenheim (2000, 461).24 Struktur und Komplexität des Follikels sind dabei ebenfalls zu berücksichtigen, wie Prum (1999, 292) bemerkt: „… jede vollständige Theorie über den Ursprung der Federn muss nicht nur die Struktur der Feder selbst erklären, sondern auch die Struktur und Komplexität des Follikels.“ Ähnlich argumentieren Maderson et al. (2000, 705): „Der Follikel und die Feder evolvierten zusammen, sie entwickeln sich zusammen und der Follikel ist eine genauso wichtige Erfindung wie die Feder.“ Die interstrukturellen Zusammenhänge der Federorganisation bzw. -funktionalität müssen letztlich im Kontext der Gesamtentwicklung des Organismus gedeutet werden: „Darüber hinaus muss die Erklärung der Körperhülle der Vögel und der Federn in einer ähnlich ganzheitlichen Weise im Zusammenhang der Evolution des ganzen Tieres betrachtet werden“ (Bock 2000, 480).
Die Anordnung der Federn auf der Körperoberfläche bei Vögeln kann selektiv kaum vorteilhaft sein ohne Steuerung durch ein effektives Kontroll- und Regulationssystem. Ebenso wenig können Federn als individuelle Strukturen evolviert sein, sondern müssen von Anfang an als gesamte Körperbedeckung entstanden sein, da der skeleto-muskuläre Apparat nicht als isolierte Einheit funktionieren kann, so Homberger & De Silva (2000, 561). Aus demselben Grund – so diese beiden Autoren – konnten Federn nicht in isolierten Reihen von Flugfedern an den Vorderextremitäten oder am Schwanz von postulierten reptilartigen Vogelvorläufern entstanden sein, wie manchmal angenommen wird.25
Für die Flugfähigkeit sind weiterhin Körperkontrolle und gute Manövrierbarkeit sehr wichtig. Diese Fähigkeiten müssen bereits von Anfang an vorhanden gewesen sein, dazu gehört auch die Landung (Caple et al. 1983, 456, 466).26Die vorstehenden Ausführungen machen deutlich, dass die Strukturen, die als Mindestvoraussetzungen für die Flugfähigkeit erforderlich sind, vielfach und über mehrere Ebenen vernetzt sind. Man kann daher im Falle von Vogelfedern (inklusive ihrer Verankerung und Steuerung) von „nichtreduzierbarer Komplexität“ sprechen.
Synorganisation spielt beim Bau der Feder somit eine wichtige Rolle: „Unser Verständnis der funktionalen Biologie der Feder und ihrer Rolle in der Evolution des Vogelflugs erfordert implizit eine Integration der Befunde über ihre faserige Mikrostruktur und die besser erkennbare grobe Anatomie“ (Lingham-Soliar & Murugan 2013, 2). „Biologische Materialien wie die Feder sind hierarchisch organisiert und detailliert konstruiert, so dass das endgültige Design es schwierig macht, die einzelnen Aspekte ihrer Funktionen vom Ganzen zu trennen. Es ist eine Art Fluidität zwischen den Funktionen und ihren Teilen (Multifunktionalität), die eine größere Anpassungsfähigkeit an verschiedene Stressfaktoren ermöglicht“ (Lingham-Soliar 2014, 324).
|
|
Zusammenfassung des Erklärungsbedarfs und Fazit
Eine Erklärung der Entstehung der Vogelfeder muss folgende Aspekte berücksichtigen:
- Die Vogelfeder ist eine komplexe Neuheit aus mehreren Innovationen: Follikel (Art des Federwachstums), röhrenförmige Federanlage, Federstruktur mit Haken- und Bogenstrahlen.
- Federmaterial, Federbau, Federkleid, Steuerungssysteme u. a. sind vielfach verschachtelt synorganisiert, wobei ganz verschiedene Ebenen aufeinander abgestimmt sein müssen: vom passenden Baumaterial über die Form von Konturfedern, deren Feinbau, Verankerung, Bewegung, Steuerung, Koordination im Verbund des ganzen Federkleids, der Skelettanatomie bis zum Verhalten und der Gehirnorganisation. Daher ist eine Diskussion zur Evolution der Vogelfeder ohne Berücksichtigung des Integuments unrealistisch (Bock 2000, 479).
- Die Struktur und Komplexität des Follikels, auch im Zusammenhang mit ihrer ontogenetischen Entwicklung (Prum 1999, 292);
- die Entstehung der Gefiederpflege mit den dafür erforderlichen Pflegemitteln und Verhaltensweisen;
- die komplexen und koordinierten Vorgänge bei der Mauser, gerade auch in Bezug auf die perfekte zeitliche Koordination mit saisonal bedingten Verhaltensweisen der Vögel (Brutzeit, Migration).
Man kann im Sinne der Evolutionstheorie argumentieren, dass für die Flugfähigkeit nicht alle genannten Fähigkeiten ausgebildet sein müssen. Sofern es dafür aus der rezenten Vogelwelt keine Analogie gibt, wäre das aber spekulativ. Aber selbst wenn dies zugunsten evolutionärer Ansätze zugestanden wird, ändert das nichts daran, dass auch die einzelnen Strukturen hochkomplex sind und nach allem unserem Wissen auch sein müssen, um zu funktionieren. So ist beispielsweise die Mauser ein hochkomplexer, koordinierter Vorgang, der nur in seiner Gesamtheit funktional und selektierbar ist und als nichtreduzierbares Subsystem gelten kann.
Es wird somit deutlich, dass die einzelnen Module und Ebenen (vom Baumaterial bis zum Verhalten) nicht isoliert voneinander verstanden werden und auch nicht isoliert entstanden sein können. In Summe haben wir mit dem Federkleid eine vielfach verschachtelte und vernetzte Gesamtorganisation vor uns, die insgesamt in Bezug auf die Flugfähigkeit nichtreduzierbar komplex erscheint und ein klares Design-Indiz darstellt. Denn nach allem unserem Wissen wird eine top-down-Organisation benötigt, für welche einerseits natürliche Prozesse weit überfordert sind, die andererseits ein typisches Kennzeichen für Planung ist.
|
Literatur
- Bergman J (2003)
- The evolution of feathers: a major problem for Darwinism. TJ 17, 33-41.
- Bock WJ (2000)
- Explanatory history of the origin of feathers. Amer. Zool. 40, 478-485.
- Burckhardt C, Fölsch DW & Scheifele U (1979)
- Das Gefieder des Huhnes. Animal Management / Tierhaltung Volume 9.
- Caple G, Balda RP & Willis WR (1983)
- The physics of leaping animals and the evolution of preflight. Am. Nat. 121, 455-476.
- Homberger DG & De Silva KN (2000)
- Functional microanatomy of the feather-bearing integument: implications for the evolution of birds and avian flight. Amer. Zool. 40, 553-574.
- Junker R & Scherer S (Hg, 2013)
- Evolution – ein kritisches Lehrbuch. Gießen, 7. Aufl.
- Kämpfe L (2003)
- Federentstehung und Vogelflug – neue evolutionsbiologische Gesichtspunkte. Praxis Nat. – Biol. in der Schule 6, 40-46.
- Kamphuis A (2008)
- Vögel: Die große Bild-Enzyklopädie. München.
- Lin CM, Jiang TX, Widelitz RB & Chuong CM (2006)
- Molecular signaling in feather morphogenesis. Curr. Opin. Cell Biol. 18, 730-741.
- Lingham-Soliar T (2014)
- Feather structure, biomechanics and biomimetics: the incredible lightness of being. J. Ornithol. 155, 323-336.
- Lingham-Soliar T & Murugan N (2013)
- A new helical crossed-fibre structure of β-keratin in flight feathers and its biomechanical implications. PLoS ONE 8(6): e65849.
- Lucas AM & Stettenheim PR (1972)
- Avian anatomy. Integument. Agriculture Handbook 362, U.S. Department of Agriculture, Washington, D.C.
- Maderson PFA, Homberger D et al. (2000)
- Symposium on evolutionary origin of feathers: Panel discussion. Amer. Zool. 40, 695-706.
- Martin LD (2008)
- Origins of avian flight – a new perspective. Oryctos 7, 45-54.
- Nachtigall W (1985)
- Warum die Vögel fliegen. Hamburg – Zürich.
- Peters DS (1994)
- Die Entstehung der Vögel. Verändern die jüngsten Fossilfunde das Modell? In: Morphologie & Evolution. Symposien zum 175jährigen Jubiläum der Senckenbergischen Naturforschenden Gesellschaft. Frankfurt/M, S. 403-423.
- Peters DS (2001)
- Probleme der frühen Vogelevolution. I. Die Sache mit den Federn. Nat. Mus. 131, 387-401.
- Proctor NS & Lynch PJ (1993)
- Manual of ornithology. Avian structure & function. Yale Univ. Press.
- Prum RO (1999)
- Development and evolutionary origin of feathers. J. Exp. Zool. 285, 291-306.
- Prum RO & Brush AH (2002)
- The evolutionary origin and diversification of feathers. Quart. Rev. Biol. 77, 261-295.
- Prum RO & Brush AH (2003)
- Zuerst kam die Feder. Spektr. Wiss. 10/03, 32-41.
- Stettenheim PR (2000)
- The integumentary morphology of modern birds – an overview. Amer. Zool. 40, 461-477.
|
|
|  |


