Studium Integrale Journal 18. Jahrgang / Heft 2 - Oktober 2011 Seite 106 - 115
|
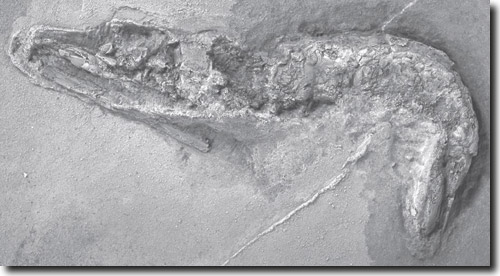
Über einen hervorragend erhaltenen Fund einer fossilen Garnele berichten Wissenschaftler von der Kent State University von Ohio. Das acht Zentimeter große Tier, das nach seinem Entdecker Royal Mapes Aciculopoda mapesi benannt wurde, zeigt sogar Details der Muskulatur im Hinterleib. Eine (mineralische) Erhaltung von Muskeln kommt nur sehr selten vor und wird auf Phosphatisierung zurückgeführt. Das Muskelgewebe blieb vermutlich durch eine Kombination aus säurehaltigem Wasser und Sauerstoffmangel so lange erhalten, bis es phosphatisiert war. Die Einbettung muss maximal binnen Wochen, also ziemlich schnell, erfolgt sein. Sogar einzelne Muskelstränge sind erkennbar. Das Fossil weist große Ähnlichkeiten mit heutigen Garnelen auf, auch wenn es in eine neue Familie (Aciculopodidae) der Zehnfußkrebse (Dekapoda) gestellt wurde (Feldmann & Schweitzer 2010). Es besitzt die charakteristische Körperform der Überordnung Eucarida, die zu den Höheren Krebsen (Malacostraca) gehört. Garnelen haben sich also seit Beginn ihrer Fossildokumentation kaum verändert; auch der neue Fund zeigt den typischen langgestreckten, etwa zylindrischen und seitlich leicht zusammengedrückten Körper. Besonders überraschend aber ist das geologische Alter des Fundes. Das Fossil wurde im oberdevonischen Woodford-Schiefer bei Ada, Oklahoma, entdeckt, der auf etwa 360 Millionen Jahre datiert wird. Zuvor galt eine 245 Millionen Jahre alte Garnele aus Madagaskar als ältestes Fossil dieser Gruppe. Die Ur-Garnele aus Oklahoma ist also mehr als 100 Millionen Jahre älter und gehört zu den beiden ältesten Dekapoden-Fossilien. Das ist erstaunlich, denn damit muss angenommen werden, dass Garnelen über einen riesigen Zeitraum in geologisch nicht überlieferten Lebensräumen existiert haben. Grund für das Fehlen weiterer Fossilbelege könnte der Lebensraum des Tieres in der Tiefsee sein. In Pressemeldungen wird kommentiert, dass die versteinerte Garnele einen wichtigen Schritt bei der Erforschung der Entwicklung der Zehnfußkrebse darstelle. Doch der neue Fund dokumentiert gerade nicht, wie der Garnelen-Bauplan entstanden sein könnte, sondern weitet den Bereich der Fossilüberlieferung aus und ist ein Beleg für „Stasis“, das Stehenbleiben auf einer bestimmten Organisationsstufe. Fragen zur Evolution werden damit gerade nicht geklärt, es werden nur neue Rahmenbedingungen für die Suche nach möglichen Vorfahrenstadien gesetzt. R. Junker [Feldmann RM & Schweitzer CE (2010) The Oldest Shrimp (Devonian: Famennian) and Remarkable Preservation of Soft Tissue. J. Crust. Biol. 30, 629-635.] |
Ein zweiter und dritter Blick auf einen Untersuchungsgegenstand ist oft lohnend! Ein baltischer Bernstein (ca. 37 x 16 x 6 mm; Eozän; Alter: ca. 40 Millionen radiometrische Jahre), in dem eine Insektenlarve an einer Spinne sitzend eingeschlossen ist, wurde erstmals von Janzen (2002) abgebildet und als Käferlarve auf einer nicht identifizierten Spinne beschrieben. Wunderlich (2004) schreibt zu der Abbildung: „Eine unbestimmte Käfer-Larve 1 mm, greift möglicherweise eine unbestimmte Jungspinne an oder sie parasitiert sie; Körper-Länge 2,1 mm. Baltischer Bernstein, Slg. J. Janzen. Es könnte sich allerdings auch um Phoresie handeln.“ (Phoresie [gr.: pherein = tragen] ist eine vorübergehende Transportgemeinschaft.) Bei der Spinne handelt es sich nach J. Wunderlichs jüngster Bestimmung möglicherweise um eine Jugendform, d. h. noch nicht geschlechtsreife, weibliche Angehörige der Sackspinnen (Clubionidae). Eine genauere Bestimmung ist aufgrund unzugänglicher Merkmale (Geschlechtsöffnung, Epigyne) nicht möglich. Michael Ohl, ein Insektenspezialist und Kurator am Museum für Naturkunde in Berlin, hat jetzt die auf der Spinne aufsitzende Insektenlarve genauer charakterisiert und als erstes Larvenstadium eines Vertreters von Fanghaften (Mantispidae) identifiziert. Die Fanghaften gehören zur Ordnung der Netzflügler (Neuroptera) und die erwachsenen Tiere sehen – wie die lateinische Bezeichnung erahnen lässt – den Fangschrecken oder Gottesanbeterinnen (Mantodea) auffallend ähnlich. Wie bei Mantodea ist das erste Brustsegment verlängert und daran sitzen die charakteristischen Fangbeine, deren Ober- und Unterschenkel mit Dornen bewehrt sind. Von den Larven der Fanghafte (Mantispidae) ist bekannt, dass sie sich ausschließlich in den Eiablagen von Spinnen entwickeln, wo sie sich von den Eiern ernähren. Im ersten Larvenstadium sind die Insekten ausgesprochen mobil und bewegen sich schnell über den Untergrund auf der Suche nach Spinnen-Eigelegen. Entweder suchen die Larven direkt nach den Eiablagen oder sie besteigen eine Spinne und verbleiben dort, bis es zur Eiablage kommt. Während des Aufenthalts auf der Spinne ernähren sich die Fanghaftlarven von der Hämolymphe (Körperflüssigkeit, die die Funktion des Blutes übernimmt) und zeigen sich sehr anpassungsfähig, d. h. sie haben ein umfangreiches Verhaltensrepertoire je nach Lebensweise des von ihnen gewählten Zwischenwirts. In späteren Stadien erscheinen die Larven eher wie Maden, die sich, wenn der Nahrungsvorrat an Spinneneiern aufgebraucht ist, nach dem 3. Larvenstadium verpuppen. Im Fossilbefund sind bisher kaum mehr als 10 Mantispidae beschrieben (früher Jura bis Miozän), davon nur 4 als Bernsteininklusen. Aufgrund molekularbiologischer Analysen wird vermutet, dass die Insektengruppe, der die Fanghafte zugeordnet werden, bereits in der frühen Trias entstanden ist. Bisher sind keine Fanghafte in Baltischem Bernstein beschrieben worden. Die von Ohl beschriebene Larve stellt also den ersten Vertreter dieser Insektenfamilie dar. Der Autor vermutet aufgrund dieses Fundes, dass die komplexen Entwicklungsstadien der Fanghaftlarven auf Spinnen bzw. deren Eigelegen bereits in den ursprünglichen Formen der Mantispinae vorhanden war. Ob alle bis heute beobachteten und beschriebenen Verhaltensweisen bereits so alt sind, kann aufgrund dieses Fundes aber nicht geklärt werden. Interessanterweise betont Ohl gleich eingangs in seinem Beitrag, dass die auffallende Ähnlichkeit von Mantispidae und Mantodea ein exzellentes Beispiel für Konvergenz darstellt. Die auffälligen und typischen Fangbeine sollen aufgrund morphologischer und molekularer cladistischer Analysen innerhalb der Netzflügler (Neuroptera) mindestens zweimal unabhängig voneinander entstanden sein. Damit liegt hier ein auffälliges Beispiel für Konvergenz vor, das sich gegen evolutionäre Erklärungen sperrt. Dass sich diese typischen Fangbeine mehrfach bei den Netzflüglern und dann auch noch bei den Fangschrecken entwickeln, sollte nicht nur festgestellt werden, sondern auch als Herausforderung für evolutionäre Erklärungen wahrgenommen werden. All dies zeigt, wie auch Ohl in seiner Arbeit betont, wie wenig die Geschichte und die Verwandtschaftsbeziehungen bei den Mantispidae bisher verstanden sind. Man darf mit ihm zusammen darauf gespannt sein, was zukünftige – in Arbeit befindliche – Studien an neuen Erkenntnissen liefern werden. H. Binder [Janzen J-W (2002) Arthropods in Baltic amber. Ampyx Verlag, Halle; Ohl M (2011) Abroad a spider – a complex developement strategy fossilized in amber. Naturwissenschaften DOI 10.1007/s00114-011-0783-2; Wunderlich J (2004) Fossil spiders in amber and copal. Beitr. Araneol. 3A, 559 Abb. 605.] |
Vor mehr als 150 Jahren beschrieben Koch & Berendt (1854) eine Spinne in Baltischem Bernstein aus der Sammlung von Berendt. Diese wird im Museum für Naturkunde in Berlin aufbewahrt. Zwei Exemplare wurden in verschiedenen Arbeiten erwähnt, dabei aber unterschiedlich beschrieben. Die Bernsteinstücke waren unter den Lagerbedingungen stark gedunkelt (Oxidation), so dass die Einschlüsse durch lichtmikroskopische Untersuchung nur eingeschränkt erkennbar waren und somit nur unzureichend charakterisiert werden konnten. Die von Koch & Berendt (1854) als Ocypete crassipes bezeichneten fossilen Spinnen (in einem Fall handelt es sich um eine Exuvie1) gaben Anlass zu kontroverser Diskussion. Denn die Spinnen gehören zu den Riesenkrabbenspinnen (Sparassidae) mit einer bekannten maximalen Körpergröße bei erwachsenen Tieren von 12,8-31,3 mm (ohne Beine) und sollten sich leicht aus dem Harz befreien können. (Aus Laos sind Sparassidae beschrieben mit einer Beinspannweite von über 30 cm.) Um die vermutlich ältesten bekannten Fossilien (Baltischer Bernstein, Eozän, nach radiometrischer Datierung 44-49 Millionen Jahre alt) von Riesenkrabbenspinnen aus der Sammlung Berendt zu überprüfen und abzusichern haben Dunlop et al. (2011) die Proben mittels einer speziellen Methode der Computer-Tomographie (CT) untersucht. Dabei wurde durch besondere Vorkehrungen der Phasenkontrast erhöht und damit auch noch feinste anatomische Strukturen sichtbar gemacht. Die Autoren erwähnen in ihrer Arbeit nicht nur z. T. kuriose Ereignisse aus der Geschichte dieser Probenstücke (wie z. B. Verwechslung der zugehörigen Beschriftungen, die die Genehmigung für eine solche Untersuchung überhaupt erst ermöglicht hatten); sie demonstrieren auch die Eignung der CT als zerstörungsfreie Analysenmethode für Bernsteinproben, nachdem Befürchtungen vorgebracht worden waren, dass durch die entsprechenden Bestrahlungen das Material leidet und zerstört wird. Durch die spezielle CT, mit der die beiden Stücke in einem Labor der Universität in Manchester (UK) untersucht wurden, konnten die Autoren z. B. die Zahnbewehrten Kieferklauen (Cheliceren) und die Anordnung der Augen sehr detailliert darstellen. Auch von den Spinnenbeinen konnten Aufnahmen erzeugt werden, die strukturelle Besonderheiten, ihre Gelenke und Behaarung sowie die Tarsalklauen mit ihren charakteristischen Häkchen zeigen. Anhand dieser charakteristischen Details konnten Dunlop et al. bestätigen, dass es sich bei den Museumsstücken um den bislang ältesten Beleg für Riesenkrabbenspinnen (Sparassidae) handelt, und damit die ursprünglich behauptete Zuordnung auch anhand moderner taxonomischer Kriterien belegen. Sie ordnen die fossile Spinne als Eusparassus crassipes in das System der Spinnen ein. Mit dieser Arbeit haben die Autoren gezeigt, dass CT mit verstärktem Phasenkontrast sehr gut geeignet ist, um undurchsichtige und gealterte Stücke fossilen Harzes und die darin eingeschlossenen Fossilien zu untersuchen. In diesem Fall konnten die Inklusen (Spinnen bzw. Exuvie) mit sehr hoher Auflösung abgebildet werden. Diese historischen Stücke stellen die ältesten Belege für die Spinnenfamilie Sparassidae und der Unterfamilie Eusparassinae dar, nämlich aus dem Eozän.
Die ältesten fossilen Belege (mit einem radiometrischen Alter von 44 bis 49 Millionen Jahren) weisen also die charakteristischen Merkmale der Eusparassinae bereits auf und sind somit für die Systematik der Spinnen ein eindrucksvoller Hinweis, dass diese Merkmale über lange Zeiträume stabil ausgeprägt werden. Eine weiterhin spannende Frage im Blick auf die ökologischen Randbedingungen, die zur Zeit der Einbettung dieser Spinnen in das Harz geherrscht haben, bzw. auf die Eigenschaften des frischen Harzes ergibt sich daraus, dass eine robuste Spinne mit einer Beinlänge von bis zu 17 mm darin gefangen worden ist. War das Harz so schnell fließend, dass sie nicht entkommen konnte? War die Harzmenge so groß, dass sie sich nicht daraus befreien konnte? Es gibt noch viele spannende Fragen, deren Antworten vielleicht ja schon in den Lagern von Museen zur „Wiederentdeckung“ bereitliegen. H. Binder [Dunlop JA, Penney D, Dalüge N, Jäger P, McNeil A, Bradley RS, Withers PJ & Preziosi RF (2011) Computed tomography recovers data from historical amber: an example from huntsman spiders. Naturwiss. 98, 519-527; Koch CL & Berendt GC (1854) Die im Bernstein befindlichen Crustaceen, Myriapoden, Arachniden und Apteren der Vorwelt. In: Berendt GC (ed) Die im Bernstein befindlichen organischen Reste der Vorwelt; 1 (II). Nicholai, Berlin, 1-124.] Anmerkung
|
Mit der Hilfe von Tausenden von Freiwilligen hat eine Gruppe von Wissenschaftlern Daten über die Farbmuster der Gehäuse von Hain-Bänderschnecken (Cepaea nemoralis) in ganz Europa gesammelt (Silvertown et al. 2011). Anhand dieser Daten wollten sie testen, welche Auswirkungen die momentane Klimaerwärmung auf diese Schnecken hat.
Die Zeichnungen der Gehäuse dieser sehr farbenfrohen Schnecke weisen Unterschiede sowohl in Farbe als auch in der Anzahl der Streifen auf; entsprechend unterscheidet man verschiedene sogenannte Morphen. Es gibt gelbe, braune und rosafarbene; und die Streifenzahl kann zwischen null und fünf Streifen schwanken (vgl. Abb. 1). Die Forscher interessierten sich für die Häufigkeit der Allele (Zustände der Gene) dieser verschiedenen Morphen. Ihre Hypothese war, dass Selektion in jüngster Zeit hellere Schneckenhäuser bevorzugen würde. Diese reflektieren mehr Licht und halten die Schnecke daher kühler in dem sich erwärmenden Klima als dunklere Gehäuse. Um dies zu testen, legten sie aus verschiedenen Quellen zwei Datensätze an (einen historischen und einen aktuellen, d. h. vor und nach dem Jahr 2000), in denen sie Information über die Farbe, die Anzahl der Streifen, den Lebensraum und die geographischen Rahmenbedingungen von über einer halben Million Schnecken sammelten. Mit Hilfe dieser Daten untersuchten sie, wie sich die Häufigkeit der Phänotypen „Gelb“, „Keine Streifen“ und „Ein Streifen“ in verschiedenen Lebensräumen (Wald, Hecke, Feld, Düne) im Verlauf der vergangenen fünfzig Jahre verändert hat. Aufgrund ihrer Hypothese erwarteten sie, dass „Gelb“ und „Keine Streifen“ in der gesamten Population häufiger anzutreffen sein würde. Die Resultate überraschten. Sowohl gelbe als auch nicht gestreifte Schnecken wurden generell weniger, die Häufigkeit der gestreiften hingegen stieg an. Außerdem wurde bemerkt, dass dort, wo die Temperatur am stärksten anstieg, die Häufigkeit der Schnecken mit dem Merkmal „Ein Streifen“ (deren Gehäuse aufgrund von Reflexion mehr kühlen sollten als die Gehäuse mit mehr Streifen) entgegen der Erwartung gesunken ist. Die Forscher mussten daher ihre Hypothese verwerfen. Die Autoren sind sich nicht sicher, warum dies so ist, schlugen aber mehrere Möglichkeiten vor, die die Resultate erklären könnten. Es könnte zum Beispiel sein, dass der Jagddruck auf die hellen Schnecken größer ist als der Selektionsdruck durch Erwärmung. Denn gelbe Schnecken wären wahrscheinlich für Fressfeinde meistens leichter zu entdecken als dunklere. Außerdem kann es gut sein, dass die Schnecken, anstatt sich genetisch zu verändern, ihr Verhalten ändern und mehr Zeit im Schatten verbringen. Diese letzte Hypothese wird durch die Beobachtung unterstützt, dass die relative Häufigkeit der gelben Schnecken dort am größten ist, wo es am wenigsten Schatten gibt (in den Dünen), aber dann stetig abnimmt je mehr Schatten in dem jeweiligen Lebensraum zur Verfügung steht (über Feld und Hecke bis hin zum Wald). Diese Studie zeigt einmal mehr sowohl den großen Einfluss, den der Lebensraum auf eine Population von Lebewesen hat, als auch die Bedeutung der Polyvalenz in der Anpassung an diesen Raum. Die verschiedenen beobachtbaren Morphen von C. nemoralis sind ein Beispiel für die genetische Variabilität (das ist ein Aspekt von Polyvalenz) eines Grundtyps. Nach dem Konzept der Grundtypenbiologie (Junker & Scherer 2006) ermöglicht diese Polyvalenz den Lebewesen, sich an veränderliche Umgebungen anzupassen, so wie es hier besonders deutlich mit dem gelben Phänotyp passiert ist. Dies kann offensichtlich sehr schnell passieren; in dieser Studie wurden lediglich 15-20 Generationen von Schnecken untersucht. Damit ist die Polyvalenz ein wichtiges Kennzeichen von Lebewesen, das sie auch bei sich schnell ändernden Bedingungen vor dem Aussterben schützt. Dass eine solche Anpassung in kurzer Zeit möglich ist, ist durch diese Studie einmal mehr gezeigt worden. Interessant ist aber auch ein anderer Aspekt: Es zeigt sich, wie schwierig es ist, phänotypische Änderungen mit Selektionsbedingungen in einen eindeutigen Zusammenhang zu bringen. Die Verflechtungen der Lebewesen mit ihrer belebten und unbelebten Umwelt sind viel zu komplex für einfache Modelle. D. Vedder [Junker R & Scherer S (2006) Evolution – Ein kritisches Lehrbuch. Gießen, 6. Auflage; Silvertown J, Cook L, Cameron R, Dodd M, McConway et al. (2011) Citizen Science Reveals Unexpected Continental-Scale Evolutionary Change in a Model Organism. PloS ONE 6(4): e18927.doi:10.137/journal.pone. 0018927] |
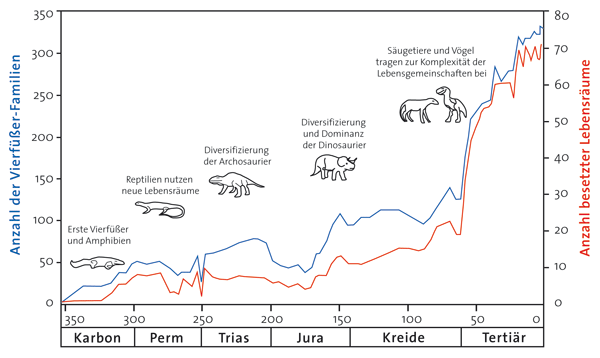
Im Jahr 1996 veröffentlichte der Paläontologe Michael Benton eine umfangreiche Analyse über die Verbreitung der Tetrapodengruppen (Vierbeiner) in verschiedenen geologischen Zeiten. Er gelangte zur Schlussfolgerung, dass eine Verdrängung von Arten durch Konkurrenz wahrscheinlich eine geringe Rolle in der Geschichte der Vierbeiner gespielt habe. Stattdessen habe es hauptsächlich eher eine Ausbreitung in freie Lebensräume gegeben. Dies widerspreche der Vorstellung Darwins, dass Evolution vor allem durch einen Verdrängungswettbewerb voranschreite. Im vergangenen Jahr veröffentlichte Benton mit Mitarbeitern eine detailliertere Folgestudie (Sahney et al. 2010), aufgrund der die Forscher diese Interpretation bestätigt sehen. Sie untersuchten den Zusammenhang zwischen der taxonomischen und der ökologischen Vielfalt in einem globalen Maßstab und stellten eine 97%ige Korrelation zwischen der weltweiten Vielfalt der Tetrapoden und ökologischen Lebensweisen (ecological modes) fest. Das heißt, mit der (phasenweise exponentiellen) Zunahme der Tetrapoden-Taxa ging (fast immer) parallel eine entsprechende Zunahme der ökologischen Vielfalt einher (Abb. 1). Neben dieser Korrelation stützen die Autoren ihre Ansicht, dass vor allem Einwandern in freie Räume und weniger Konkurrenz und Verdrängung die Verbreitung der Tetrapoden ermöglichte, unter anderem auch darauf, dass die Tetrapoden nur etwa ein Drittel der verfügbaren ökologischen Nischen einnehmen.
Die Autoren bestreiten nicht, dass Konkurrenz auf niedrigem taxonomischem Level (z. B. zwischen Arten) eine wichtige Rolle spielt. Für die größeren Veränderungen der Vielfalt von Tetrapodengruppen halten sie jedoch ihre hier vorgestellten Argumente für ausschlaggebend. Es geht also nicht darum, ob Darwin in Bezug auf Konkurrenz falsch lag, wie manche Pressetexte suggerierten, sondern wie weit dieses Konzept trägt und wo seine Grenzen liegen. Die Interpretation von Sahney et al. (2010) erfuhr auch Kritik, so z. B. durch den Hinweis, dass Dinosaurier und frühe Säugetiere 60 Millionen Jahre lang in Koexistenz gelebt hätten, mithin die Dinosaurier gleichsam wettbewerbsmäßig überlegen gewesen seien, was eben doch auf Konkurrenz hinweise. Erst nach ihrem Aussterben hätten sich dann die Säugetiere stark ausgebreitet. Andererseits zeigt gerade die Koexistenz, dass es keine Verdrängung durch Konkurrenz gegeben hat. In einer weiteren aktuellen Studie untersuchten Vamosi & Vamosi (2010) bei Angiospermen (bedecktsamige Blütenpflanzen, die größte Pflanzengruppe), ob es einen Zusammenhang zwischen der Artenanzahl von Angiospermen-Familien und ihrem mutmaßlichen phylogenetischen Alter gibt. Dabei konnten sie entgegen den evolutionär begründeten Erwartungen keine Korrelation feststellen. Vielmehr besteht ein Zusammenhang mit der Größe des besiedelten geographischen Gebietes und (an zweiter Stelle) mit Merkmalen von Blüten und Früchten. Die Korrelation mit der Größe des Besiedlungsgebietes passt ebenfalls eher zur Kolonisierungs- als zur Konkurrenzhypothese und unterstützt damit die These von Sahney et al. (2010). Wenn die Verteilung fossiler Taxa eher auf Eroberung freier Lebensräume als auf Konkurrenz und Wettbewerb beruhen, stellt sich zum einen die Frage nach dem Motor der Veränderungen. Einwandern in freie Lebensräume bedeutet geringere Selektionsdrücke als Existenz unter harten Konkurrenzbedingungen. Jedoch wird man hier kaum scharf argumentieren können. Zum anderen aber könnte ein Besiedlungsszenario auch so interpretiert werden, dass Arten, die bereits in (nicht fossil überlieferten) Lebensräumen bzw. begrenzten Rückzugsräumen existierten, sich in freie Ökosysteme ausgebreitet haben und damit auch mit höherer Wahrscheinlichkeit fossil überliefert wurden. Für die gesamte Fossilabfolge wird diese Erklärung nicht ausreichen, möglicherweise aber für einzelne geologische Systeme; insbesondere wenn man geologischen Hinweisen folgt, die auf sehr viel kürzere Zeiträume hindeuten. Exponentielles Wachstum an Formenvielfalt könnte vor allem Ausdruck von Einwanderungen sein, also ökologische und weniger evolutionäre Gründe haben. R. Junker [Benton MJ (1996) Testing the roles of competition and expansion in tetrapod evolution. Proc. R. Soc. B 263, 641-646. (doi:10.1098/rspb.1996.0096); Sahney S, Benton MJ & Ferry PA (2010) Links between global taxonomic diversity, ecological diversity and the expansion of vertebrates on land. Biology Letters 23, 544-547. (doi: 10.1098/rsbl.2009. 1024); Vamosi JC & Vamosi SM (2010) Key innovations within a geographical context in flowering plants: towards resolving Darwin’s abominable mystery. Ecology Letters 13, 1270-1279. (doi: 10.1111/j.1461-0248. 2010.01521.x); Vgl. auch den Blogbeitrag von David Tyler unter http://www.arn.org/blogs/ index.php/literature/2010/09/06/explanations_of_vertebrate_diver-sity] |
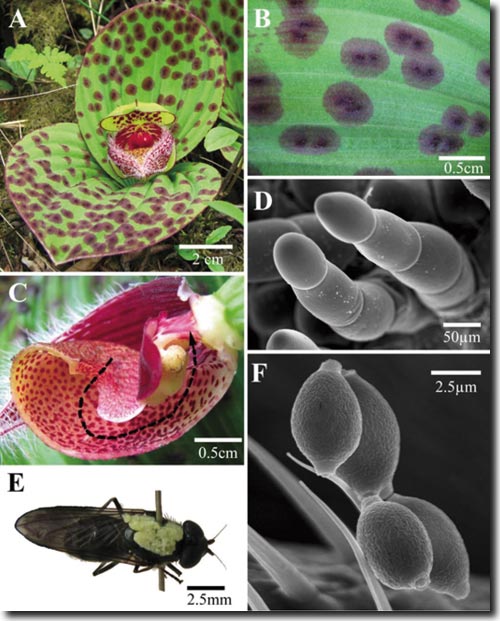
In letzter Zeit wird zunehmend deutlich, dass mehr Blütenpflanzen als bisher angenommen ihre Bestäuber durch Vortäuschung von Nahrung anlocken, ohne ihnen die für Bestäuber übliche „Belohnung“ zu gewähren (vgl. im gleichen Heft den Artikel über die Bestäubung in der Gattung Aronstab und das nachstehende Streiflicht über die Situation bei der Orchideen-Art Epipactis veratrifolia). Bei der artenreichen Familie der Orchideen sind Insektentäuschblumen häufig. Man schätzt, dass etwa ein Drittel aller Orchideen-Arten, also etwa 6000-10000 Arten, ihre Bestäuber täuschen. Dabei trifft man immer wieder auf ausgeklügelte und überraschende Situationen. Als Lockmittel dienen neben visuellen Reizen vor allem die für die Orientierung von Insekten so wichtigen Duftstoffe. So haben jetzt chinesische und amerikanische Wissenschaftler (Ren et al. 2011) festgestellt, dass bei der mit dem heimischen Frauenschuh verwandten Art Cypripedium fargesii aus Südwestchina (Abb. 1 A) eine sonst bei Blütenpflanzen als Bestäuber nicht bekannte Fliegenart in besonderer Weise angelockt wird, um ohne Gegenleistung die Bestäubung durchzuführen. Die entsprechende, zu den Tummel- oder Pilzfliegen (Platypezidae) zählende Art aus der Gattung Agathomyia (Abb. 1 E) ernährt sich wie die meisten Vertreter dieser Familie von Mikropilzen, die parasitisch auf Laubblättern leben, hier speziell einem Schimmelpilz der Gattung Cladosporium. Die Orchidee imitiert die Struktur und die typische Schwarzfärbung dieses Pilzes durch schwarz behaarte Flecken auf der Blattoberfläche (Abb. 1 B, D) bei gleichzeitiger Abgabe von Duftmolekülen durch die Blüte. Diese erinnern an den Geruch faulender Blätter und sind in sehr ähnlicher Form auch von Kulturen des Pilzes bekannt. Merkwürdigerweise sehen außerdem die mehrzelligen Haare in den Blattflecken (Abb. 1 D) sehr ähnlich aus wie die Konidiosporen abgebenden Pilzfäden von Cladosporium (Abb. 1 F). Die auf diese Weise angelockten Fliegen gelangen schließlich in die Blüte. Genauso wie die Bestäuber unseres heimischen Frauenschuhs (Cypripedium calceolus) müssen sich die Fliegen, um wieder herauszukommen, durch die sackförmige Ausstülpung der Blüte hindurch zwängen und führen dabei die Bestäubung durch. Nektar als potentielle „Belohnung“ wird nicht geboten, und der Pollen landet in Form der für Orchideen typischen Gesamtpakete (Pollinien) auf dem Rücken des Insektes (Abb. 1 E). Im vorliegenden Fall täuscht also eine Blütenpflanze einen phytopathogenen Pilz vor, um Insekten für die eigene Fortpflanzung zu missbrauchen. An dieser Stelle sei aber erwähnt, dass es in der Natur auch den umgekehrten Fall gibt, nämlich dass phytopathogene Pilze Blüten vortäuschen (Pseudoblüten) und dadurch Insekten anlocken, die für die Fortpflanzung des Pilzes sorgen. Dies wurde z. B. ausführlich für die Beziehung zwischen dem Rostpilz Puccinia arrhenatheri und dessen Wirtspflanze Berberis vulgaris (Sauerdorn, Berberitze) beschrieben (Naef et al. 2002). Hier werden die Insekten durch visuelle und olfaktorische (Geruchs-) Reize (Mimikry) angelockt, bekommen aber auch eine nektarhaltige Flüssigkeit zur „Belohnung“. In allen Fällen handelt es sich um „geniale“ Strategien zur Aufrechterhaltung der Fortpflanzung. H. Kutzelnigg [Naef A, Roy BA, Kaiser R & Honegger R (2002) Insect-mediated reproduction of systemic infections by Puccinia arrhenatheri on Berberis vulgaris (Berberidaceae). New Phytologist 154, 717-730. http://ceeb.uoregon.edu/faculty_pages/Roy/papers/35.pdf; Ren Z-X, Li D-Z, Bernhardt P & Wang H (2011) Flowers of Cypripedium fargesii (Orchidaceae) fool flat-footed flies (Platypezidae) by faking fungus-infected foliage. Proc. Natl.Acad.Sci. 108, 7478-7480.] | |||||
Ganz anders gelagert als im vorstehend geschilderten Fall der Frauenschuh-Art Cypripedium fargesii, aber ebenso eindrucksvoll ist die Situation bei einer weiteren Orchidee, der Stendelwurz-Art Epipactis veratrifolia (= Germerblättrige Stendelwurz). Bei dieser in Vorderasien heimischen, bis ca. 1,50 m hohen Art werden die Bestäuber – es sind hier verschiedene Schwebfliegen-Arten (Familie Syrphidae) – in wieder völlig anderer Weise angelockt (Stökl et al. 2011). Die hier zahlreich vorhandenen Blüten (Abb. 1) täuschen nämlich durch warzenartige Strukturen, vor allem aber durch die Abgabe flüchtiger chemischer Verbindungen einen Blattlausbefall vor. Das abgegebene Duftstoffgemisch mit α- und β-Pinen, β-Myrcen und α-Phellandren als Hauptkomponenten entspricht nämlich sehr genau den Alarmstoffen (Alarm-Pheromonen) verschiedener Blattläuse. Die Schwebfliegen (darunter die auch bei uns häufige Hain-Schwebfliege Episyrphus balteatus) werden von diesem Geruch stark angezogen, weil die Weibchen ihre Eier bevorzugt in die Nähe von Blattlaus-Ansammlungen legen. Denn die schlüpfenden Larven ernähren sich räuberisch von Blattläusen. Im Zuge der Eiablage bekommen die Schwebfliegen-Weibchen nach der für Orchideen typischen Art die Pollenpakete (Pollinien) angeheftet, was dann bei einem weiteren Blütenbesuch zur Bestäubung führt. Auch Männchen halten sich zwecks Begattung in der Nähe der Blüten auf und beteiligen sich gelegentlich an der Bestäubung. Die Schwebfliegen erhalten in den Blüten zwar eine kleine Portion Nektar. Dennoch handelt es sich insgesamt um eine Täuschung, weil zum einen durch die Abgabe der Alarmstoffe falsche Tatsachen vorgespiegelt werden, und zum anderen weil keine Blattläuse vorhanden sind und somit die aus den Eiern schlüpfenden Larven dem Tode geweiht sind. Man vermutet, dass etwa 10% aller Orchideen-Arten durch Vortäuschung eines Eiablageplatzes ihre Bestäuber täuschen. Die Situation bei Epipactis veratrifolia ist dabei der erste beschriebene Fall einer Anlockung durch Imitation der Alarmstoffe von Blattläusen. In der etwa 30-60 Arten umfassenden Gattung Epipactis gibt es eine große Vielfalt an Bestäubungssystemen, z. B. durch Wespen oder Hummeln. Blütendüfte wurden chemisch bisher nur bei solchen Arten untersucht, die vor allem durch Wespen bestäubt werden. Sie unterscheiden sich grundlegend von den Duftstoffen von E. veratrifolia. H. Kutzelnigg [Stökl J, Brodmann J, Dafni A, Ayasse M & Hansson BS (2011) Smells like aphids: orchid flowers mimic aphid alarm pheromones to attract hoverflies for pollination. Proc. R. Soc. B 279, 1216-1222.] | |||||
Der Birkenspanner (Biston betularia) ist wohl eines der berühmtesten Lehrbuchbeispiele für beobachtete Evolution und speziell für das Wirken der Selektion. Seine dunkle Form (als carbonaria bezeichnet) war ab Mitte des 19. Jahrunderts beobachtet worden. Sie nahm im Zuge der Industrialisierung und der damit verbundenen Umweltverschmutzung erheblich zu und löste die hellen Formen (typica) zeitweise in einigen Regionen weitgehend ab. Nach 1950 drehte sich der Trend wieder um. Ende der 1990er Jahre gab es einige Diskussionen um diesen Falter. Denn es stellte sich heraus, dass die Ursachen für die Ausbreitung der dunklen Form und die Selektionsbedingungen komplexer sein müssen als zuvor angenommen (Majerus 1998, Coyne 1998, vgl. Stud. Int. Journal 6 (1999), 97; www.wort-und-wissen.de/sij/sij62/sij62-s.html). Unklar war bisher nach wie vor die genetische Basis für den Unterschied zwischen dem Wildtyp (helle Form) und der dunklen Form. Waren Mutationen, die zur carbonaria-Form führten, erst in jüngerer Zeit aufgetreten, wieviele Mutationen liegen zugrunde und sind sie mehrfach unabhängig aufgetreten? Eine von van’t Hof et al. (2011) publizierte Genanalyse kam nun zum Ergebnis, dass alle dunklen Formen dasselbe Allel des betreffenden Gens besitzen und dass es erst in jüngerer Zeit entstanden ist und starker Selektion ausgesetzt war. Damit scheint geklärt zu sein, dass die dunkle Form nur einmal entstanden ist und sich unter den sich verändernden Umweltbedingungen des 19. Jahrhunderts in England und auf die Isle of Man schnell ausbreiten konnte. R. Junker [Coyne JA (1998) Not black and white. Nature 396, 35-36; Majerus MEN (1998) Melanism: Evolution in action. Oxford; Van’t Hof AE, Edmonds N, Dalíkova M, Marec F & Saccheri IJ (2011) Industrial Melanism in British Peppered Moths Has a Singular and Recent Mutational Origin. Science 332, 958-960.] |
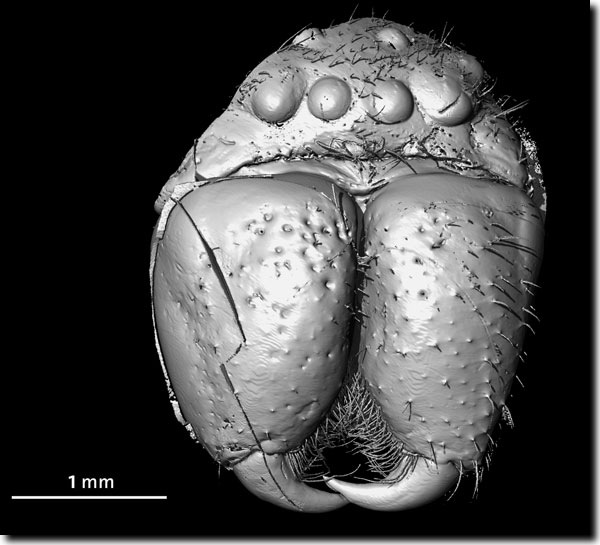
Die Evolution der Augen ist ein vieldiskutiertes Thema in der Evolutionsbiologie und unter Evolutionskritikern (vgl. Ullrich et al. 2005). Dabei stehen Vergleiche verschiedener Augentypen heutiger Arten und theoretische Überlegungen über Neuerwerb einzelner Bestandteile komplexer Augen z. B. durch sogenannte Kooptionen im Vordergrund. Welchen Beitrag aber liefern Fossilfunde? Hier ist weitgehend Fehlanzeige zu verzeichnen. „Der Fossilbericht war bis jetzt unzureichend, um Einblicke in die frühe Evolution der Augen während der anfänglichen Radiation vieler Tiergruppen zu ermöglichen, die als kambrische Explosion bekannt ist“ (Lee et al. 2011, 631). Außer von Trilobiten-Augen sei kaum etwas über die Details des optischen Designs der kambrischen Tierwelt, aus der die ältesten fossil erhaltenen Augen stammen, bekannt, trotz z. T. sehr guter fossiler Erhaltung. Nun berichten Lee et al. (2011) von sehr gut erhaltenen fossilen Augen aus dem frühen Kambrium des Emu Bay-Schiefers in Australien (auf 515 Millionen Jahre datiert), die sie als „sehr hochentwickelte“ (highly advanced) Komplexaugen (Facettenaugen) charakterisieren. Sie sind in fein geschichtetem Tonstein eingebettet. Es handelt sich um mehrere isolierte Augen, die aus jeweils über 3.000 ommatidischen Linsen in dichter und in ausgeprägt sechseckiger Anordnung bestehen, was die kompakteste und effizienteste Anordnung darstellt. (Als Ommatidien werden die Einzelelemente von zusammengesetzten Augen bezeichnet, wie sie in der heutigen Tierwelt bei Gliederfüßern vorkommen.) Der Durchmesser der Augen beträgt 7-9 mm. Die Forscher ordnen die Augen aufgrund ihrer Größe einem Gliederfüßer zu, der vermutlich räuberisch lebte und unter schwachen Lichtverhältnissen sehen konnte. Eine genauere Zuordnung ist nicht möglich. Die Augen sind komplexer als fossile Augen zeitgleich existierender Trilobiten und so hochentwickelt wie die Augen vieler heute lebender Formen. Die Augen besitzen im Zentrum große ommatidische Linsen, die eine besonders lichtempfindliche „helle Zone“ („bright zone“ oder „acute zone“) bilden, während die Linsen in den Randbereichen kleiner sind. Diese Spezialisierungen seien für viele moderne Taxa charakteristisch. Die Wissenschaftler schließen aus der Anordnung der verschiedenen Linsengrößen, dass die Augen auch im Lebenszustand flach waren. Die extrem reguläre Anordnung der Linsen übertrifft sogar die Anordnung bei manchen heute lebenden Formen wie dem Pfeilschwanz Limulus. Damit besaßen einige der ältesten Gliederfüßer optische Systeme, die denen heutiger Formen glichen. R. Junker [Lee MSY, Jago JB, García-Bellido DC, Edgecombe GD, Gehling JG & Paterson JR (2011) Modern optics in exceptionally preserved eyes of Early Cambrian arthropods from Australia. Nature 474, 631-634; Ullrich H, Winkler N & Junker R (2005) Zankapfel Auge. Ein Paradebeispiel für „Intelligent Design“ in der Kritik. Stud. Int. J. 13, 3-14.] |
Stanley S. Miller gilt als der Chemiker, der als Pionier Mitte des 20. Jahrhunderts erste experimentelle Untersuchungen zur Chemie der Lebensentstehung angestellt hat. Er veröffentlichte 1953 eine Aufsehen erregende und bis heute unverändert populäre Arbeit, in der er in eindrucksvoller Weise demonstrierte, dass man im Labor aus einem einfachen Gasgemisch Aminosäuren, die Bausteine für Proteine synthetisieren kann. Miller, der im Mai 2007 im Alter von 77 Jahren gestorben ist, hat sich lebenslang mit den Fragen des chemischen Ursprungs von Leben beschäftigt. Auch wenn er hinsichtlich einer naturwissenschaftlichen Klärung dieser Frage optimistisch war, so hat er doch gleichzeitig nüchtern und kritisch die bislang vorgeschlagenen Szenarien analysiert und kommentiert. Johnson et al. (2008) identifizierten in Proben 22 Aminosäuren (neben vielen nicht proteinogenen Aminosäuren (die in heutigen Proteinen nicht vorkommen) auch Glycin, Alanin, Serin, Asparaginsäure, Valin, Glutaminsäure und Phenylalanin) und fünf Aminoverbindungen. Sie zeigten damit, dass die klassischen Ursuppen-Experimente hinsichtlich der Vielfalt an gebildeten Aminosäuren ergiebiger waren, als Miller dies erkennen konnte, aber auch, dass viele Verbindungen entstehen, die hinsichtlich biologischer Funktionen keine erkennbare Bedeutung haben oder für die Bildung von Peptiden hinderlich sind. Bada und ein Team von Mitarbeitern veröffentlichten nun erneut Analysenergebnisse von Proben aus Millers Simulationsexperimenten, die dieser 1958 durchgeführt, aber nie veröffentlicht hatte (Parker et al. 2011). Aus einem Gasgemisch bestehend aus Methan (CH4), Ammoniak (NH3), Schwefelwasserstoff (H2S) und Kohlendioxid (CO2) hatte Miller in einer modifizierten Miller-Apparatur Simulationsexperimente unter besonderer Berücksichtigung von Vulkanausbrüchen durchgeführt. (Im Vergleich zur Originalapparatur war eine konisch zulaufende Glasspitze als Düse vor dem eigentlichen Reaktionsraum eingebaut: Glaskolben mit eingeschmolzenen Elektroden.) Insgesamt konnten Parker et al. 23 Aminosäuren und vier Aminoverbindungen identifizieren. Dabei enthielten 6 der Aminosäuren Schwefel (S) ebenso wie eine der Aminoverbindungen. Bei den nachgewiesenen 6 S-haltigen Aminosäuren handelt es sich nicht um solche, die am Aufbau von Proteinen beteiligt sind. Die Aminosäuren waren aus 2 bis 6 C-Atomen aufgebaut und alle, sowohl die proteinogenen als auch die anderen Aminosäuren waren racemisch, d. h., die spiegelbildlichen Isomere waren innerhalb der Messgenauigkeit im Verhältnis 1:1 vorhanden (D/L = 1 ± 10%). Diesen Befund werten die Autoren auch als Hinweis darauf, dass die Proben während der Lagerung nicht oder höchstens in Spuren verunreinigt wurden (zumindest nicht durch Verunreinigungen biologischen Ursprungs). Cystein, die einzige S-enthaltende Aminosäure, die in Proteinen vorkommt, konnte nicht nachgewiesen werden, oxidierte verwandte Verbindungen dagegen sehr wohl. Die Autoren spekulieren deshalb darüber, dass Cystein möglicherweise ursprünglich bei diesen Experimenten produziert worden war, aber während der Lagerung (ohne Schutz vor Oxidation) chemisch verändert worden ist. Die Autoren diskutieren die mögliche Bedeutung von Vulkanen, die in der Erdgeschichte zumindest lokal und für gewisse Zeit reduzierende Gasmischungen einschließlich Schwefelwasserstoff produziert haben könnten. Im Vergleich zu den anderen klassischen Simulationsexperimenten von Miller ist die Vielfalt an Aminosäuren größer (die proteinogenen Aminosäuren Threonin, Leucin und Isoleucin werden dort nicht beschrieben). Die Ausbeute an Aminosäuren ist in dem Experiment mit Schwefelwasserstoff (H2S) – vor allem bei den nicht proteinogenen Aminosäuren – im Vergleich zu den anderen Experimenten größer. Parker et al. (2011) vergleichen die Verteilung der Aminosäuren und deren relative Häufigkeit in den von ihnen untersuchten Proben mit Analysendaten von kohligen Chondriten, der häufigsten Klasse von Meteoriten, die – wie die Bezeichnung andeutet – C-haltige Verbindungen aufweisen. Solche Vergleiche hatte bereits Miller (Wolman et al. 1972) angestellt. Die zumindest teilweise vorhandene Ähnlichkeit nehmen die Autoren zum Anlass über vergleichbare Synthesebedingungen im interstellaren Raum bzw. an den Herkunftsorten der Meteoriten und in den Simulationsexperimenten, sowie über die Bedeutung der Gegenwart von H2S bei der Synthese von Aminosäuren zu spekulieren. Mit dieser späten Analyse, Auswertung und Publikation von Millers Simulationsexperimenten ist dokumentiert, dass Stanley Miller dabei erstmals die Synthese von S-haltigen Aminosäuren gelungen ist. H. Binder [Binder H (2003) Miller-Experimente zur Chemie der Lebensentstehung – 50 Jahre danach. Stud. Int. J. 10, 65-73; Johnson AP, Cleaves HJ, Dworkin JP, Glavin DP, Lazcano A & Bada JL (2008) The Miller vulcanic spark discharge experiment. Science 322, 404; Junker R & Scherer S (2006) Evolution – ein kritisches Lehrbuch. Gießen; Miller SL (1955) Production of some organic compounds under possible primitive earth conditions. J. Am. Chem. Soc. 77, 2351-2361; Parker ET, Cleaves HJ, Dworkin JP, Glavin DP, Callahan M, Aubrey A, Lazcano A & Bada JL (2011) Primordial synthesis of amines and amino acids in a 1958 Miller H2S-rich spark discharge experiment. Proc. Nat. Acad. Sci. USA; doi/10.1073/pnas.1019191108; Wolman Y, Haverland WJ, Miller SL (1972) Nonprotein amino acids from spark discharges and their comparison with the Murchison meteorite amino acids. Proc. Natl. Acad. Sci. USA 69, 809-811.] |

|