|
Entstehung des Flugapparats
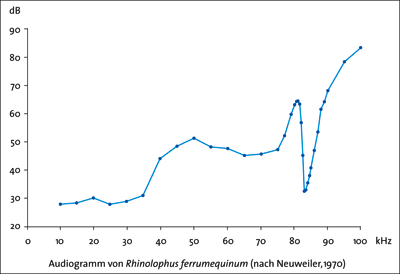 |
| Abb 3: Hörschwelle einer Großen Hufeisennase (Rhinolophus ferrumequinum). In der Vertikalen ist angegeben, ab welcher Lautstärke (in Dezibel) der Schall von der Fledermaus wahrgenommen werden kann. Bei 82-83 kHz ist ein schmales Band großer Empfindlichkeit. Die Fledermäuse stoßen Rufe in etwas geringerer Frequenz aus, so dass bei Annäherung an eine Beute aufgrund des Dopplereffekts das Echo die Frequenz des empfindlichsten Bereichs trifft. (Nach Dietz et al. 2007) |
 |
| Abb 4: Anordnung von Muskeln und elastischen Bändern bei der Rauhhautfledermaus Pipistrellus nathusii. (Nach Krapp 2004) |
|
Die Fledermäuse bilden die einzige Gruppe unter den Säugetieren, die aktiv fliegen kann. Die Mittelhand- und die Fingerknochen außer dem Daumen sind verlängert und dienen als Gerüst für die weitgehend oder völlig unbehaarte Flughaut (Patagium, im Bereich der Finger als Dactylopatagium bezeichnet). Die Flughaut verläuft nach vorne von den ebenfalls verlängerten Armknochen weiter zum Hals (Propatagium, Vorderflughaut). Sie erstreckt sich weiter an der Körperseite entlang bis zu den Beinen (Plagiopatagium, Seitenflughaut) und zwischen den Beinen (Uropatagium, Schwanzflughaut) (Abb. 1). Zum Stützen und Spannen der Schwanzflughaut besitzen viele Arten eine besondere Knochenspange, den Sporn (Calcar) an der Ferse.
Die Flughaut ist dünn, sehr elastisch, dennoch reißfest und von feinen Muskelfasern, elastischen Bändern, Nerven und Blutgefäßen durchzogen (Abb. 4). „Diese Blutgefäße zeigen ein selbständiges, rhythmisches Pulsieren; dadurch ist die gleichmäßige Blutversorgung auch der entferntesten Teile der Flughaut gesichert“ (Eisentraut 1968a, 89), außerdem wird dadurch eine schnelle Wundheilung ermöglicht. Für die sehr empfindliche Flughaut verwenden die Fledermäuse viel Zeit mit der Pflege. Die Flughaut darf nicht austrocknen und muss geschmeidig gehalten werden; die Fledermäuse reiben sie mit einem Drüsensekret aus der Mundfalte ein. Ihre weichen und durch viele Muskeln justierbaren Flügel erlauben den Fledermäusen ein hervorragende Manövrierbarkeit und Wendigkeit (Neuweiler 1993, 27). Die Flügeloberfläche ist mit tastempfindlichen Rezeptoren ausgestattet, sogenannten Merkel-Zellen, die es zwar auch bei anderen Säugetieren gibt, bei den Fledermäusen aber noch ein feines Härchen im Zentrum besitzen, wodurch sie noch empfindlicher sind und Informationen über Luftbewegungen im Flügelbereich sammeln können, um den Flug entsprechend anpassen zu können. Weitere Rezeptoren bei Arten, die mit den Flügeln Insekten fangen, können die Dehnung der Flugmembran ermitteln und dadurch Informationen über den Beuteerwerb gewinnen.
Die Fingerknochen sind biegsam und sehr viel flexibler als bei anderen Säugetieren und im Querschnitt abgeflacht. Angesichts der Leichtbauweise besteht die Gefahr eines Durchschlagens der dünnen Flughaut. Dies wird dadurch verhindert, dass die verlängerten Finger als „Spannsparren eingesetzt werden und die Flughaut an der Körperwand und den Beinen festgezurrt wird.“ Die Fledermäuse können die Wölbung ihrer Flügel verändern und auf diese Weise den Flugbewegungen anpassen (Neuweiler 1993, 8). In der Ruhestellung werden die Flügel wie eine Ziehharmonika zusammengefaltet an den Körper angelegt und die Flughaut zieht sich zusammen und wird verkleinert.
Mit der Flugfähigkeit sind zahlreiche Besonderheiten des Bauplans gekoppelt, die das Achsenskelett, den Schultergürtel, die seitliche Orientierung der Hinterbeine und die kopfstehende, hängende Ruhestellung betreffen (Gunnell & Simmons 2005, 210). „Der besonderen Beanspruchung der Vordergliedmaßen beim Fliegen entspricht die kräftig entwickelte Flugmuskulatur und ein sehr fester Schultergürtel. Der große Brustmuskel sitzt am verknöcherten Brustbein an, das in der Mittellinie ähnlich wie bei den Vögeln einen flachen Kamm hat. Das Schultergelenk ist ein kompliziert gebautes Kugelgelenk, das die rudernden Bewegungen des Flügels gestattet. Ellenbogen, Hand- und Fingergelenke sind Scharniergelenke, die zusammen mit der entsprechenden Muskulatur der ausgespannten Flügelfläche festen Halt geben“ (Eisentraut 1968a, 89). „Fast alle Besonderheiten der Fledermäuse hängen in irgendeiner Weise mit ihrer Fähigkeit zu fliegen zusammen“ (Dietz et al. 2007, 28).
Wenn man die Entstehungsweise des Flugapparats aufklären möchte, muss man sich die vorgenannten Details und Zusammenhänge vor Augen halten. Die wichtigsten Indizien, die zur Klärung beitragen können, stammen aus der Paläontologie und der Vergleichenden Entwicklungsgenetik.
Fossilien
 |
Alle fossilen Formen besitzen bereits
alle Kennzeichen des aktiven Flugs. |
 |
|
Über die Entstehung des Flugapparats geben Fossilien nahezu keine Auskunft. Die ältesten fossil überlieferten Fledermäuse erscheinen plötzlich und vielfältig im unteren Eozän (vor ca. 52 Millionen Jahren nach radiometrischen Datierungen), so zum Beispiel Icaronycteris index aus der Green-River-Formation in Wyoming (Abb. 5). Bei dieser Gattung wird lediglich eine kleine Klaue auf dem Zeigefinger als Rest von bodenlebenden Vorfahren interpretiert (Simmons 2008, 99), die Beinproportionen werden als etwas primitiver eingestuft als bei anderen Fledermäusen, die Finger besitzen rudimentäre Endphalangen und es fehlt ein Calcar (Gunnell & Simmons 2005, 214). Simmons (2008, 99) stellt dennoch fest: „Ironically, however, perhaps the most remarkable thing about Icaronycteris is just how much this ancient beast resembles extant bats.“ Auch alle anderen untereozänen Formen wie Archaeonycteris (Abb. 6) oder Palaeochiropteryx aus Europa besitzen bereits alle Kennzeichen des aktiven Flugs (Sears et al. 2006, 6581; Gunnell & Simmons 2005, 209) und der Fledermaus-Bauplan ist seit seinem fossilen Erscheinen weitgehend konstant geblieben (Neuweiler 2003, 249).
 |
| Abb 5: Icaronycteris index aus dem Eozän von Wyoming (Wikipedia) |
 |
| Abb 6: Archaeonycteris trigonodon aus der Grube Messel bei Darmstadt (Wikipedia) |
|
Aufgrund molekularer Daten werden große Lücken in der Fossilüberlieferung der meisten Fledermauslinien vermutet. Von der mutmaßlichen Zeit der Existenz der Linien der Fledermausfamilien sind durchschnittlich 73% ohne Fossilbeleg (Teeling et al. 2005, 583). Teeling et al. sehen darin einen möglichen Grund für das Fehlen von Übergangsformen.
Bereits aus dem mittleren Eozän sind viele Fledermausarten überliefert, die modernen Familien zugeordnet werden können (Gunnell & Simmons 2005, 209, 217) und die ältesten fossil bekannten Fledermäuse überlappen zeitlich mit modernen Familien (Teeling et al. 2005, 582). Einige fossile Familien können heutigen Gruppen nicht zugeordnet werden. Die aus der weltberühmten mitteleozänen Grube Messel bei Darmstadt bekannten sieben Fledermausarten besaßen so unterschiedliche Flügelformen, „dass man sie den Flug- und Jagdbiotopen heutiger Fledermausgesellschaften zuordnen kann. Auch die Variationsbreite von verschiedenen aerodynamisch wichtigen Parametern wie Flügelform und Flächenbelastung deckte bereits quantitativ die Variationsbreite heutiger Fledermäuse ab“ (Habersetzer et al. 2008, 246). Wie Icaronycteris besaßen die Messelfossilien „nahezu alle einzigartigen Merkmale heutiger Fledermäuse“ (Gunnell & Simmons 2005, 214).
Die Fledermäuse waren im Eozän auch geographisch weit verbreitet, es sind Funde aus Europa, Nordamerika, Nordafrika, Indien und Australien bekannt, die überall plötzlich auftauchen (Gunnell & Simmons 2005, 216; Smith et al. 2007, 1003; Teeling et al. 2005, 582). Bis 1998 waren 24 Gattungen aus dem Eozän bekannt (Simmons & Geisler 1998, 5), davon acht aus dem Untereozän; viele heutige Mikrochiropterenlinien können bis zum Mittel- und Obereozän verfolgt werden (Denzinger et al. 2004, 311). Der geographische Ursprung der Fledermäuse ist unbekannt (Smith et al. 2007, 1003), ebenso besteht Unklarheit über die nächsten Verwandten der Fledermäuse (Simmons 2008, 103). Simmons (2005; 2008, 101) spricht daher von einer „Big-bang-Evolution“, die auch durch Daten über DNA-Sequenzen gefordert werde. Dieses Phänomen tritt auch bei anderen Säugetiergruppen auf.
Lediglich die erst vor wenigen Jahren entdeckte fossile Gattung Onychonycteris („Krallen-Fledermaus“, Abb. 7) besaß Merkmale, die auf mögliche evolutive Vorfahren hinweisen könnten, nämlich Klauen an allen fünf Fingern sowie relativ kurze Vorderarme und relativ lange Hinterfüße im Vergleich zu anderen Fledermäusen (Simmons et al. 2008). Dennoch besteht kein Zweifel, dass auch Onychonycteris aktiv über lange Strecken fliegen konnte, wie der robuste Knochen- und Gelenkaufbau des Flügelskeletts, der versteifte Brustkorb und ein breiter Brustbeinkiel (Ansatz für die kräftige Brustmuskulatur) zeigen. Eine zusätzliche Flughaut zwischen Hinterbeinen und Schwanzwirbelsäule hat vielleicht den möglichen Nachteil kleiner Flügel ausgeglichen. Dazu half ein langer Calcar, der oben erwähnte Knochensporn, der die Hautmembran der Schwanzflughaut aufspannte. Ein solcher fehlt bei der phylogenetisch nahe stehenden Gattung Icaronycteris und der Messeler Gattung Archaeonycteris (Habersetzer et al. 2008, 250; Simmons 2008, 100); in dieser Hinsicht ist Onychonycteris also eher „modern“. Unter den heute lebenden Fledermäusen besitzen die Mausschwanz-Fledermäuse (Rhinopomatidae) ähnlich kurze und breite Flügel wie Onychonycteris. Sie haben einen ungewöhnlichen Gleit-Flatter-Flug, indem sie den Schlagflug durch kurze Gleitphasen unterbrechen (Simmons 2008, 100).
Auch Onychonycteris ist also eine voll ausgebildete Fledermaus und ein „beträchtlicher Umfang an evolutionären Veränderungen trennt Onychonycteris und andere Fledermäuse von ihren bodenlebenden Vorfahren“ (Simmons 2008, 103). Onychonycteris ist in denselben Schichten gefunden worden wie Icaronycteris (Simmons 2008, 100). In einer morphometrischen Analyse konnten Sears et al. (2006) zeigen, dass die Fingerproportionen des 3.-5. Fingers seit Beginn der fossilen Überlieferung der Fledermäuse unverändert sind – ein weiterer Befund, der ein plötzliches Erscheinen in der Fossilüberlieferung der Fledermäuse belegt.
Entwicklungsgenetik
Fossilfunde geben also keine nennenswerte Auskunft über die Entstehung der Fledermausflügel. Im Zeitalter der Genomforschung und der Entwicklungsbiologie stehen aber weitere Erkenntnisquellen zur Verfügung, aus denen man Hinweise auf den Ursprung von Organen zu entnehmen versucht. Die Biologen suchen nach Genen, die für die Ausbildung der betreffenden Organe benötigt werden. Dazu dienen Vergleiche entwicklungsgenetisch bedeutsamer Gene von Fledermäusen mit flügellosen Säugetieren und deren Manipulation durch Veränderungen ihrer Einflussnahme auf die Entwicklungssteuerung. Aus diesem Wissen werden dann theoretische Szenarien einer evolutiven Entstehung abgeleitet bzw. spekulativ entworfen.
Im Falle des Fledermausflügels sind die Genetiker fündig geworden. Es wurden Gene identifiziert, die bei der Verlängerung der Fingerknochen und bei der Ausbildung der Flughaut eine Rolle spielen. Die Ergebnisse werden nachfolgend skizziert.
Sears et al. (2006), Weatherbee et al. (2006) und Sears (2008) berichten über die Identifizierung von Genen, die bei der Bildung der Flughaut und der Flügelknochen beteiligt sind und schließen aus ihren Befunden, dass geringe Änderungen in der Expression von Genen (also der Nutzung ihrer Information) große Änderungen im Bau der Flügel verursacht haben könnten. Säugetiere, die im Erwachsenenzustand keine Haut zwischen den Fingern ausbilden, besitzen solche Häute im embryonalen Stadium, bevor durch programmierten Zelltod das Gewebe zwischen den Fingern zurückgebildet wird. Dabei spielt der BMP-Signalweg eine wichtige Rolle als Regulator. BMP-Gene sind in den Regionen zwischen den Fingern aktiv und verursachen den Zelltod. Bei den Fledermäusen wird jedoch im Gewebe zwischen den Fingern eine große Menge des Gens Gremlin exprimiert, ähnlich wie in den Füßen von Enten, was die Aktivität von BMP unterdrückt. Das alleine genügt jedoch nicht, um das Gewebe zwischen den Fingern zu erhalten; es muss als zweiter Inhibitor der Fibroblast-Wachstumsfaktor fgf8 („fibroblast growth factor“) dazukommen, damit das Flughautgewebe ausgebildet wird (Sears 2008, 7; Weatherbee et al. 2006; vgl. Abb. 8).
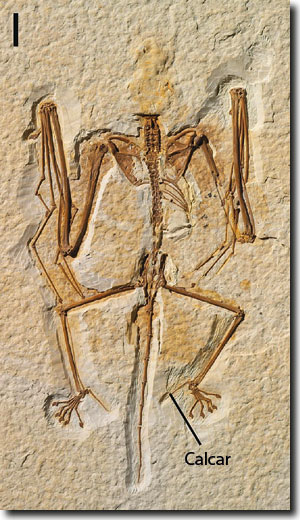 |
| Abb 7: Holotyp von Onychonycteris finneyi. Balken: 1 cm. (Aus Simmons et al. 2008, Abdruck mit freundlicher Genehmigung) |
|
Es wurden außerdem Gene identifiziert, die einen Einfluss auf die Länge der Armknochen und der Finger haben. In der Embryonalentwicklung bilden sich die Finger bei Fledermäusen und Mäusen zunächst in gleicher Größe und werden bei den Fledermäusen dann im Laufe der Ontogenese (Individualentwicklung) verlängert. Bei den Fledermäusen erfolgt die Bildung der Knorpelzellen in den Extremitätenanlagen gegenüber den Verhältnissen bei Mäusen beschleunigt, so dass mehr Knorpelzellen pro Zeiteinheit gebildet werden (Sears et al. 2006, 6585). Dabei sind viele molekulare Signalwege beteiligt (Sears 2008, 9); es wurden mehrere Gene identifiziert, die bei der Reifung der Knorpelzellen eine Rolle spielen (Sears et al. 2006, 6581). Dem BMP-Signalweg kommt auch hier eine tragende Bedeutung zu, er erscheint im Vergleich zu anderen Organismen hier deutlich ausgeprägter, d. h. BMP wird vermehrt exprimiert. Auch das Wirkungsareal von FGF8 in der Randleiste an der Spitze der Extremitätenknospe (AER) ist im Vergleich dreifach vergrößert, ein Phänomen, dessen Ursache vermutlich von anderen vorgeschalteten Genen erst ermöglicht wird. Der Zu-sammenhang mit der Verlängerung der Knochen ist hier aber noch nicht gesichert und erfordert weitere Untersuchungen (Sears 2008, 9).
Ein weiteres Erfordernis für den Flug ist die Verringerung des Gewichts. So ist die Elle bei den Fledermäusen reduziert (vgl. Abb. 9). Dafür sind nach Sears (2008, 10) mehrere Entwicklungsprozesse in drei Schritten erforderlich: Bildung und nachfolgender Verlust des Knorpels, eine geringere relative Rate der Verlängerung der Elle und Verschmelzung mit benachbarten Skelettelementen. Der Autor stellt weiter fest, dass geklärt werden müsse, ob weitere Mechanismen geltend gemacht werden müssen, um die korrespondierenden Änderungen in den umgebenden Geweben wie Muskeln und Sehnen zu ermöglichen, die für den aktiven Flug benötigt werden (S. 11).
Als weitere gut mögliche Kandidaten für die Regulation des Skelettwachstums diskutiert Sears (2008, 11) den Einfluss der Hox-Gene; hier sind zur genaueren Klärung weitere Untersuchungen erforderlich.
 |
| Abb 8: Schematische Darstellung der Unterschiede der Genexpression bei Mäusen (freie Zehen), bei Enten (Zehen mit Häuten) und bei Fledermäusen. Hohe Konzentration des Bmp-Signalwegs verursachen Zelltod zwischen den Fingern. Bei den Enten wird dies durch die Aktivität von Gremlin (Gre) verhindert, bei den Fledermäusen wirkt außerdem Fgf8 als Inhibitor. (Nach Weatherbee et al. 2006) |
|
Kritische Anmerkungen
Die deutlich ausgeprägtere Wirkung des BMP-Signalwegs scheint also eine wichtige Voraussetzung für die größere Länge der Finger der Fledermäuse zu sein. Sears (2008, 10) merkt an, dass es „nicht notwendigerweise der einzige“ Mechanismus sei. Darüber hinaus ist die Länge der Finger nur ein Parameter, der für die Flugtauglichkeit der Flügel angepasst werden muss. Denn auch die Form und die unterschiedliche Länge der einzelnen Finger müssen passend ausgebildet sein. Eingangs wurde auch erwähnt, dass die Fingerknochen besonders flexibel und abgeflacht sind, was offenbar für die Funktionsfähigkeit des Flügelgerüsts notwendig ist. Die Einschätzung von Sears (2008), dass eine kleine Änderung in der Expression von Schlüsselgenen zu großen Änderungen im Bau der Knochen und der Membran führte, erscheint vor diesem Hintergrund viel zu optimistisch und verkürzt (vgl. Kastentext). Bisher wurden in Wirklichkeit nur sehr wenige Voraussetzungen der ontogenetischen Ausbildung des Flügels geklärt. Über phylogenetische Mechanismen ist damit noch gar nichts gesagt.
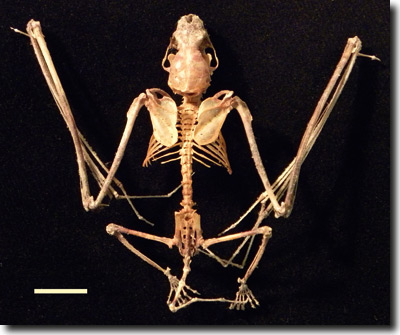 |
| Abb 9: Skelett des in Nord-amerika beheimateten Mausohrs Myotis lucifugus. Balken: 1 cm. (Wikipedia) |
|
Sears (2008, 7) schränkt an anderer Stelle selbst ein, dass die Mechanismen, die für die Bildung der meisten Teile der Flughaut verantwortlich sind, erst noch erforscht werden müssen. Schließlich können große Teile der Flughaut nicht durch das Ausbleiben des programmierten Zelltods erklärt werden, von den besonderen oben genannten unentbehrlichen Eigenschaften der Flughaut ganz zu schweigen (s. Kastentext). So merken Weatherbee et al. (2006, 15105) an, dass die molekularen Grundlagen der Regulation des Plagiopatagiums (Haut entlang des Körpers) bei gleitenden Säugetieren unbekannt sei und dass dieses Gewebe bei den Fledermäusen auf eine ganz andere Weise gebildet werde als das Dactylopatagium (Fingerflughaut). Und was die Fingerknochen betrifft, so ist es mit deren Verlängerung alleine bei weitem nicht getan. Erklärt werden müssen ja auch die spezifisch unterschiedlichen Längen der Fingerknochen, ihre Abflachung und ihre Flexibilität. Und auch all dies genügt bei weitem nicht, da beim Flug auch viele andere Organe und Gewebe mitwirken. Diese Problematik wird im Abschnitt „Was ist eine Erklärung?“ im zweiten Teil des Artikels weiter vertieft werden.
Wir können kurz zusammenfassen:
1. Es ist nur ein sehr kleiner Teil der genetischen Grundlagen der Ontogenese der Flügel bekannt. Mit der Identifizierung von Genen, die für die Ausprägung eines Organs notwendig sind, liegen Befunde zum Verständnis der ontogenetischen Entwicklung vor, die bei evolutionären Betrachtungen berücksichtigt werden müssen.
2. Selbst wenn diese Grundlagen sehr viel weiter aufgeklärt wären, wüsste man noch nichts über den Modus und die Mechanismen einer evolutiven Entstehung im Verlauf der hypothetischen Stammesgeschichte, da man aus Mechanismen der Ontogenese nicht auf Abläufe während der hypothetischen Phylogenese schließen kann.
3. Ebenso ungeklärt bleiben bei den diskutierten spekulativen Erklärungsansätzen die Wechselbeziehungen zwischen den postulierten ontogenetischen Veränderungen im Verlauf der Fledermausevolution und der sie bedingenden bzw. kanalisierenden Selektionsdrücke.
|
• elastische, mit Muskeln und Bändern ausgestattete, reißfeste und reparierbare Flughaut; • Flughäute nicht nur zwischen den Fingern, sondern auch am Körper entlang; • biegsame, abgeflachte Fingerknochen; • Form und Länge der Knochen für jeden Finger passend gestaltet; • zusammenklappbare Flughaut; • reduzierte Elle und weitere Gewichtsersparnisse; • Spezielle Flugmuskeln und deren Koordination; • Leichtbauweise des ganzen Skeletts; • spezielle Fähigkeiten des Gehirns und des Verhaltens
|
|


