 |
|
von Reinhard Junker
Studium Integrale Journal
16. Jahrgang / Heft 2 - November 2009
Seite 74 - 80
|
 |
Zusammenfassung: In den ersten beiden Artikelfolgen über Evo-Devo (Junker 2008; 2009) wurde erläutert, dass Evo-Devo-Forscher natürliche Auslese (Selektion) auf der Basis genetischer (erblicher) Variabilität für nicht ausreichend halten, um die Entstehung evolutiver Neuheiten (Makroevolution) zu erklären. Das motiviert die Suche nach Quellen von Variation, die in der Synthetischen Evolutionstheorie wenig oder keine Beachtung finden. Im zweiten Beitrag wurde die Neuverschaltung von Genen diskutiert. Als weitere Quelle für evolutionär Neues wird genetische Akkommodation* betrachtet: Extreme Umwelten sollen die Flexibilität der Organismen über das normale Spektrum umweltbedingter Veränderungsmöglichkeiten (= Modifikationen*) hinaus erhöhen. Versteckte Fähigkeiten, auf Umweltstress mit veränderten Eigenschaften zu reagieren (= kryptische Variation*), sollen Rohmaterial für Evolution bereithalten. Durch genetische Assimilation* kann die Veränderung des Erscheinungsbildes (= Phänotyp*) im Erbgut verankert werden (sog. Baldwin-Effekt*).
Evolution soll also durch eine Erhöhung der Umweltsensitivität* erfolgen, das ist eine erhöhte Anpassungsfähigkeit bei Umweltänderung. Die Umweltsensitivität soll zunächst durch Umweltstress phänotypisch sichtbar und dann auch genetisch fixiert werden: genetische Akkommodation. Bei diesem weitgehend hypothetischen Vorgang wird also die „Antwortfähigkeit“ des Phänotyps auf Umweltänderungen gesteigert und diese zugleich auch genetisch (im Erbgut) verankert.
Zur Evolution der Erhöhung der Umweltsensitivität diskutiert Moczek (2008) folgende Idee: Lebewesen sind gegen genetische und umweltbedingte Störungen abgepuffert („Entwicklungs-Belastbarkeit“, developmental capacitance). Entwicklungsprozesse können also ein gewisses Maß an genetischer Variation abpuffern und vor Selektion schützen, weil sie sich (vorerst) nicht im Phänotyp bemerkbar macht. Es gibt somit eine Art Kanalisation* in der ontogenetischen* Entwicklung. Innerhalb der „Pufferzone“ können neue versteckte (kryptische) Variationen angesammelt werden. Wenn nun bestimmte Schwellen der verkraftbaren Umweltstörungen überschritten werden, dann kann eine Änderung phänotypisch „durchbrechen“ und dadurch in Erscheinung treten. Nun kommt es darauf an, dass diese phänotypisch sichtbar gewordene Änderung zufällig an die neue Umwelt angepasst ist und daher ausgelesen werden kann. Gleichzeitig muss aber auch eine Mutation eintreten, die diese Änderung fixiert (genetische Assimilation) und das bisherige Spektrum der Modifikationen verschiebt. Der Vorteil dieser Idee ist, dass umweltbedingte Modifikationen sich nicht wie Mutationen allmählich in einer Fortpflanzungsgemeinschaft durchsetzen müssen, sondern dass sie sofort und gleichzeitig alle ihre Mitglieder betreffen, solange der betreffende Umweltfaktor gegeben ist; sie können sich also viel schneller in einer Population ausbreiten. Allerdings muss die später notwendige genetische Änderung sich dennoch wie alle genetischen Änderungen in der Population ausbreiten.
In diesem dritten Teil der Artikelserie über Evo-Devo sollen genetische Assimilation, genetische Akkommodation und „Entwicklungs-Belastbarkeit“ vorgestellt werden. Es soll diskutiert werden, ob der Mechanismus der genetischen Akkommodation realistischerweise zu evolutiven Neuheiten führen kann und ob er als Quelle für schnelle mikroevolutive Veränderungen in Frage kommt.
| |
|
Viele Jahrzehnte lang wurde die Synthetische Evolutionstheorie von den meisten Biologen für ausreichend angesehen, um die Entstehung evolutiver Neuheiten (Makroevolution) zu erklären. Deren Kernstück ist der Zweischritt Mutation – Selektion. Das heißt: Evolution vollzieht sich durch Änderungen des Erbguts (Genotyp), die sich in einer Änderung des Erscheinungsbildes (Phänotyp) äußern. Die neuen Phänotypen müssen sich bewähren und durch natürliche Selektion gefördert werden. Ist dies der Fall, können sich die zugrundeliegenden Mutationen in der Population (Fortpflanzungsgemeinschaft) durchsetzen. Evolutive Änderungen verlaufen also in der Richtung Genotyp -› Phänotyp.
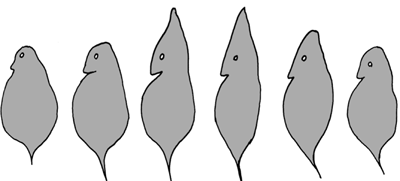 |
| Abb. 1: Cyclomorphose beim Flohkrebs Daphnia. Die Entwicklung des „Helms“ (mittlere Individuen) wird durch die Anwesenheit eines Räubers ausgelöst und kann auch einfach durch bestimmte Chemikalien im Wasser provoziert werden, auch wenn der Räuber gar nicht anwesend ist. Näheres im Text. |
|
In den bisherigen beiden Folgen wurde jedoch auf Forscher hingewiesen, die dem auf Charles Darwin und Alfred R. Wallace zurückgehenden Selektionsmechanismus nur eine Teilerklärung des evolutiven Wandels zubilligen. Demnach könne mit der Synthetischen Theorie zwar Erhaltung des Bestehenden und Feinjustierung von Merkmalsausprägungen erklärt werden, nicht aber die Entstehung von Neuem. Die wesentlichen Fragen nach dessen Herkunft bleiben damit ohne Antwort. Diese Situation motiviert die Suche nach Quellen von Variation, die in der Synthetischen Evolutionstheorie kaum Beachtung gefunden haben. Im letzten Beitrag der Evo-Devo-Artikelserie (Junker 2009) wurden als mögliche neue Quellen für evolutive Neuheiten bereits Neuprogrammierungen, Mehrfachverwendung und eine Art Zweckentfremdung von Genen vorgestellt und kritisch diskutiert.
Im vorliegenden Artikel wird eine ganz andere Quelle für Veränderlichkeit der Lebewesen thematisiert. Es geht um die Plastizität (Formbarkeit) der Organismen und ihrer individuellen Entwicklung. Damit sind umweltbedingte Modifikationen gemeint, also Unterschiede im Phänotyp, die durch bestimmte Umweltreize hervorgerufen werden (Beispiele folgen weiter unten). Man spricht in diesem Zusammenhang auch von Umweltsensitivität. Das heißt: Die Lebewesen können in Abhängigkeit von den Umweltbedingungen verschiedene, bereits im Erbgut ‚schlummernde’ Entwicklungsprogramme abrufen. Dies könnte Ausgangspunkt auch für evolutionäre Veränderungen sein. Das heißt: Umgekehrt wie in der Synthetischen Evolutionstheorie sollen phänotypische Veränderungen, hervorgerufen durch Umweltänderungen (vor allem Umweltstress) eine Vorreiterrolle in der Evolution haben. Erst nach dem Auftreten von Änderungen im Phänotyp folgen dann Änderungen im Genotyp nach. Kurz: Phänotyp -› Genotyp. Das ist zweifellos gegenüber den klassischen Vorstellungen über Evolutionsmechanismen unorthodox. Die dafür vorgeschlagene Argumentationskette soll nachfolgend erläutert werden.
|
|
Modifikation, Plastizität, Flexibilität
Es ist schon lange bekannt, dass die Lebewesen ein Potenzial besitzen, je nach Umweltbedingungen unterschiedliche Merkmale oder Merkmalsausprägungen zu bilden, ohne dass eine Änderung des Erbguts zugrunde liegt. Solche nicht-erblichen Modifikationen werden von Evo-Devo-Forschern als wichtiger Schlüssel zum Verständnis von Makroevolution betrachtet. Im Folgenden wird dafür auch der Begriff „Plastizität“ verwendet. Gemeint sind damit also Änderungen in Organismen infolge von Umweltreizen (Schlichting & Smith 2002, 190). Beispielsweise bildet der Wasser-Hahnenfuß (Ranunculus aquatilis) gelappte Luftblätter mit Spaltöffnungen (für den Gasaustausch), im Wasser sind die Blätter dagegen fein zerteilt und besitzen viel Durchlüftungsgewebe. An der Wasseroberfläche entstehen Übergangsblätter. Gleiches Erbgut, aber unterschiedliche Ausprägungen (Abb. 2).
 |
| Abb. 2: Verschiedene Blattformen des Wasser-Hahnenfußes (Ranunculus aquatilis) als Beispiel für Plastizität. Im Wasser sind die Blätter zerschlitzt, an der Luft gelappt und an der Wasseroberfläche intermediär ausgeprägt. (Aus Otto Wilhelm Thomé, Flora von Deutschland, Österreich und der Schweiz 1885. Gera. Public Domain) |
|
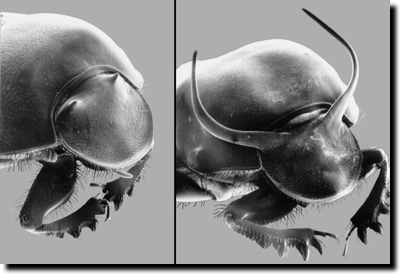 |
| Abb. 4: Polymorphismus in der Entwicklung der Hörner des männlichen Mistkäfers. Unterhalb einer bestimmten Käfergröße werden die Hörner nicht ausgebildet (Schwelleneffekt). (Aus Stern & Emlen 1999; Abdruck mit freundlicher Genehmigung von D. Emlen). |
|
Der Flohkrebs Daphnia bildet einen „Helm“ aus, wenn Räuber in der Nähe sind (Abb. 1). Die Bildung des Helms wird durch bestimmte Chemikalien ausgelöst, die von den Fressfeinden ins Wasser abgegeben werden. Der Helm behindert das Gefressenwerden, er erfordert aber Stoffwechselaufwand und verringert die Schwimmfähigkeit. Das ist vermutlich der Grund, weshalb der Helm nicht immer ausgebildet bzw. wieder zurückgebildet wird, wenn Daphnia nicht von Feinden bedroht ist.
Wenn die umweltbedingten Modifikationen graduell sind, spricht man von „Reaktionsnormen“; sind die unterschiedlichen Ausprägungen dagegen sprunghaft verschieden, handelt es sich um einen Polyphenismus („Vielgestaltigkeit“). Ein bekanntes Beispiel für Letzteres ist das Landkärtchen, ein Schmetterling, der eine Sommer- und Winterform ausbildet, je nachdem, in welcher Jahreszeit die Raupen schlüpfen. Ein weiteres Beispiel ist die Hörnerbildung des Mistkäfers Onthophagus taurus, die erst dann erfolgt, wenn eine bestimmte Körpergröße überschritten wird (Schwelleneffekt; Stern & Emlen 1999; Abb. 4).
Es ist keine neue Erkenntnis, dass die Fähigkeit vererbt wird, innerhalb gewisser Grenzen je nach Umwelteinflüssen und organismusinternen Faktoren zu variieren (Nanjundiah 2003, 247). Neu bei Evo-Devo ist, hier eine Quelle für den evolutiven Wandel zu vermuten, wie wir im Weiteren sehen werden.
|
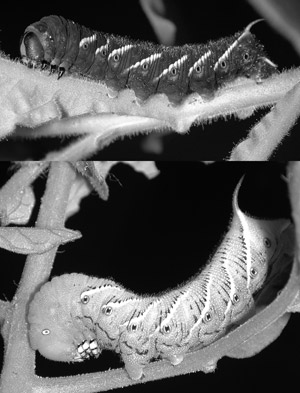 |
| Abb. 3: Die schwarze und grüne Form des Tabakschwärmers (Manduca sexta). (Foto: Yuichiro Suzuki, Abdruck mit freundlicher Genehmigung) |
|
Über die evolutive Entstehung von Polyphenismus und mithin von Umweltsensitivität (genetische Akkommodation) ist wenig bekannt. Das Problem dabei besteht darin: „Es benötigt den Erwerb von komplizierten genetischen Kontrollelementen, die die Umweltbedingungen messen und selektiv verschiedene Gengruppen in Abhängigkeit von diesen Bedingungen aktiviert“ (Myers 2006). In einer 2006 veröffentlichten Studie konnten Suzuki & Nijhout unter Laborbedingungen diesen Vorgang experimentell demonstrieren. Der Tomatenschwärmer (Manduca quinquemaculata) zeigt einen Polyphenismus: im kühleren Norden der USA sind die Raupen schwarz, was vorteilhaft für die Absorption von Sonnenlicht und damit von Wärme ist. Im wärmeren Süden sind die Raupen dagegen grün und dadurch besser getarnt (Abb. 3).
Die Autoren untersuchten nun die Raupe des nahe verwandten Tabakschwärmers (Manduca sexta), die normalerweise immer grün ist. Es existieren aber Mutanten des Tabakschwärmers mit schwarz gefärbten Raupen. Es konnte gezeigt werden, dass die Mutation zu einer Verringerung des Juvenilhormons führt, welches die Färbung der Haut reguliert. Im mutierten Stamm kamen jedoch Raupen vor, die nach einer Hitzeschockbehandlung in einem frühen Entwicklungsstadium unterschiedliche Grünfärbungen aufwiesen. Durch weitere Züchtung dieser besonderen Mutante erzielten die Wissenschaftler nach nur 13 Generationen einen Stamm, dessen Raupen ab einer bestimmten Temperaturschwelle von 28,5° C immer grün statt schwarz gefärbt waren. Die höhere Temperatur führte zur Produktion von mehr Juvenilhormon, in dessen Folge wieder die grüne Färbung auftritt. Damit war eine Umweltsensitivität (hier: Sensitivität auf unterschiedliche Temperatur) und damit die Entstehung eines Polyphenismus und mithin eine genetische Akkommodation experimentell demonstriert.
Die Experimente zeigen, dass es – als Vorstufe zur sichtbaren Variation der Färbung – eine unsichtbare Variation gibt, nämlich die Konzentration des Juvenilhormons. Ist diese niedrig, sind die Raupen schwarz gefärbt, ist sie hoch, dann sind sie grün – erst einmal unabhängig von der Temperatur. Die Variation (viel oder wenig Juvenilhormon) kann nun beim gezüchteten Tabakschwärmerstamm bzw. natürlicherweise beim Tomatenschwärmer durch die Temperatur festgelegt und fixiert werden. Es zeigte sich, dass der gezüchtete polyphene Stamm des Tabakschwärmers eine mittlere Konzentration an Juvenilhormon aufweist, die nahe beim Schwellenwert für das Umschalten von „schwarz“ auf „grün“ liegt. Durch die Temperaturerhöhung kann – wie genau ist ungeklärt – die Hormonmenge über den Schwellenwert gehoben werden, mit dem Erfolg, dass die Raupen grün statt schwarz sind (Pennisi 2006). Woher allerdings das ganze System kommt, das die Temperatursensitivität ermöglicht, ist durch diese Versuche nicht gezeigt. Offenbar wurde nur die Regulation des (an sich unsichtbaren) Hormontiters verändert (Suzuki & Nijhout 2006, 652).
Die Tatsache, dass der Tomatenschwärmer natürlicherweise polyphen ist, kann als Hinweis darauf gewertet werden, dass das Potential zur Umweltsensitivität ursprünglich in der Gattung Manduca vorliegt und beim Tabakschwärmer teilweise verlorengegangen war, jedoch unter Laborbedingungen wieder reaktiviert werden konnte. Damit wäre auch diese Umweltsensitivität nur ein Ausdruck der Polyvalenz der Stammform. Die Behauptung von Myers (2006), man habe direkt die Evolution eines „komplexen, polygenen, polyphenen Merkmals“ durch genetische Assimilation und Akkommodation im Labor beobachtet, ist durch die experimentellen Daten nicht gedeckt, wenn damit die de novo-Entstehung gemeint sein sollte.
|
Genetische Assimilation: „Einfrieren“ einer Modifikation (Baldwin-Effekt)
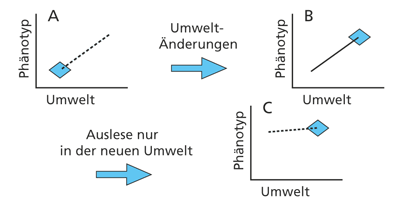 |
| Abb. 5: Schematische Darstellung der genetischen Assimilation. A: gestrichelte Linie = Modifikationsspanne. Nach einer Umweltänderung ändert sich die Merkmalsausprägung im Rahmen der Modifikationsspanne (B). Die Spanne der phänotypischen Ausprägungsmöglichkeiten verringert sich in dauerhaft veränderter Umwelt. (C). Nach Pigliucci et al. (2006) |
|
Es hat sich gezeigt, dass eine phänotypische Antwort auf eine spezifische Umwelt (also eine Modifikation) von dieser Umwelt unabhängig werden kann. Was zunächst als rein physiologische Anpassung an neue Bedingungen begann (d.h. ohne erbliche Änderung), kann offenbar bei Auftreten passender Mutationen genetisch fixiert werden. Dieser Vorgang ist schon länger als Baldwin-Effekt beschrieben worden (z. B. Nanjundiah 2003, 259; auf eine Arbeit von Baldwin [1896] zurückgehend) und wird auch als genetische Assimilation bezeichnet (vgl. Abb. 5).
Häufig wird in diesem Zusammenhang ein Experiment von Waddington als Beleg für einen solchen Vorgang angeführt. Waddington (1953) setzte sich entwickelnde Embryonen von Drosophila einem Ätherdampf aus, worauf ein Teil der Embryonen eine starke phänotypische Änderung zeigte: einen zweiten Thorax (bithorax-Form, Modifikation). Er selektierte im Folgenden kontinuierlich auf Drosophila-Fliegen mit dieser Entwicklungskapazität. Nach etwa 20 Generationen bildeten einige Fliegen die bithorax-Form ohne Äther-Behandlung. Selektion begünstigt also nach Waddington einen speziellen Entwicklungsweg, der zur Erzeugung des gewünschten Effekts führte. Schließlich wurde der Entwicklungsweg kanalisiert, daher bildete sich das Endstadium der bithorax-Form unabhängig von den Umweltbedingungen (hier: Ätherdampf) aus.
Modifikative Veränderungen des Phänotyps können demnach bei Vorliegen eines Selektionsvorteils durch nachfolgende genetische Änderungen „kopiert“ bzw. fixiert werden. Selektion entkoppelt also die Abhängigkeit der Merkmalsausprägung vom Umweltreiz (Nanjundiah 2003, 254) und begünstigt eine genetische Fixierung der ursprünglich modifikativen Veränderung (genetische Assimilation). Alternative Entwicklungswege werden somit ausgeschaltet, so dass es zu einer dauerhaften Veränderung kommt, die mit Verlust an Variabilität einhergeht.
|
|
Neue Plastizität: Genetische Akkommodation
Die Tatsache, dass die Lebewesen durch Umweltreize veränderbar sind (Modifikationen) und bestimmte Modifikationen genetisch assimiliert werden können (s. o.), ermöglicht an sich noch keine Evolution. Wie kann Neues ins Spiel kommen? Dazu müsste in einem ersten Schritt die Bandbreite der bisherigen Modifikationsmöglichkeiten verändert werden. Um zu einem evolutiven Wandel zu gelangen, stellt man sich folgendes Szenario vor: Eine stressende Umwelt (d. h. eine, die deutlich von der bisherigen Umwelt eines Lebewesens abweicht) bewirkt eine sonst nicht vorkommende Modifikation (die also das normale Spektrum von sichtbaren Modifikationen überschreitet). Wenn diese neu auftretende Modifikation auch adaptiv, also in der neuen Umweltsituation vorteilhaft ist, spricht West-Eberhard (2005, 610) von phänotypischer Akkommodation. Extreme Umwelten sollen also eine Plastizität bewirken, die über das normale Spektrum hinausgeht (vermittelt durch epigenetische* Prozesse). Versteckte Reaktionsnormen (bzw. Modifikationsspektren) könnten Rohmaterial für nachfolgende Evolution bereithalten. Damit dies zu einer dauerhaften Veränderung führt, muss eine nachträgliche genetische Fixierung und Selektion erfolgen (Nanjundiah 2003, 251): Genetische Assimilation durch Ausschalten alternativer Entwicklungswege (s. o.).
Aber auch extreme Umwelten können nur das „herausholen“, was schon an Potential in den Organismen steckt. Daher ist ein zweiter Schritt erforderlich: Die Umweltsensitivität muss durch eine genetische Veränderung erhöht werden. Neue Antwortmöglichkeiten auf Umweltänderungen (insbesondere Umweltstress) müssen evolvieren. Dieser hypothetische Vorgang wird als genetische Akkommodation bezeichnet; man könnte das als „genetisch bedingte Steigerung der ‚Antwortfähigkeit’ des Phänotyps auf Umweltänderungen“ charakterisieren. Experimentelle Belege für einen solchen Vorgang sind jedoch sehr schwach (Suzuki & Nijhout 2006; vgl. Kastentext).
Vorteil dieser „Evolutionsmethode“. Die Modifikation durch Umwelteinflüsse muss sich nicht wie Mutationen allmählich in einer Population durchsetzen, sondern betrifft alle Mitglieder einer Population, solange der betreffende Umweltfaktor gegeben ist, es gibt also eine viel schnellere Ausbreitungsmöglichkeit (gleichzeitig bei vielen Individuen – im Gegensatz zu Mutationen, die nur den einzelnen Organismus betreffen). Eine evolutive Bedeutung dieses Vorgangs wird weiter darin gesehen, dass die Plastizität und phänotypische Akkommodation nicht in zufälligen Richtungen verläuft, sondern gemäß der Maßgabe der schon vorhandenen Konstruktion. Gegenüber (ungerichteten) genetischen Veränderungen bringt die Plastizität einen „Anpassungsvorsprung“ (West-Eberhard 2005, 612). Später muss dann eine genetische Assimilation erfolgen; diese braucht aber dann doch Zeit (siehe unten).
Unterschied zur Synthetischen Theorie. Der Phänotyp ist leitend, nicht der Genotyp! „Evolution durch genetische Akkommodation unterscheidet sich daher von traditionellen Perspektiven über adaptive Evolution darin, dass Umweltstörungen, Entwicklungsprozessen und den Wechselwirkungen zwischen ihnen spezifische Rollen zugeschrieben werden, nämlich zuvor versteckte genetische Variation der Selektion verfügbar zu machen“ (Moczek 2007, 303).
|
Akkommodation: Erhöhung der Flexibilität (Anpassungsfähigkeit), Vergrößerung der Fähigkeit zu -› Modifikation (Umweltanpassung). Assimilation: Das Erblichwerden einer ursprünglich umweltbedingten -› Modifikation, so dass der betreffende Merkmalszustand immer (nicht nur bei bestimmten Umweltbedingungen) auftritt. Baldwin-Effekt: Das Ergebnis einer genetischen -› Assimilation, das Erblichwerden einer umweltbedingten Veränderung (-› Modifikation). epigenetisch: Vorgänge „oberhalb“ der genetischen Ebene. Kanalisation: Fähigkeit der Lebewesen, trotz verschiedener Umweltbedingungen ein gleichbleibendes Erscheinungsbild auszuprägen. kryptische Variation: Versteckte Ausprägungsmöglichkeiten von Merkmalen, die -› phänotypisch normalerweise nicht in Erscheinung treten. Modifikation:Umweltbedingte (durch Umweltreize ausgelöste), nicht erbliche Veränderung. Ontogenese: Individuelle Entwicklung von der befruchteten Eizelle bis zum ausgewachsenen Organismus. Phänotyp: äußeres Erscheinungsbild. Plastizität: Formbarkeit; hier vor allem die Fähigkeit, bei unterschiedlichen Umweltbedingungen verschiedene -› Phänotypen auszubilden. Polymorphismus: Vielgestaltigkeit. Population: Fortpflanzungsgemeinschaft. Umweltsensitivität: Fähigkeit, auf verschiedene Umweltbedingungen unterschiedlich zu reagieren.
|
|
|
Puffer gegen Störungen und kryptische Variation
Woher kommen die Entwicklungsmöglichkeiten, die durch Umweltreize aktiviert werden können und das Rohmaterial für Evolution bilden sollen? Dazu muss ein wenig ausgeholt werden.
Lebewesen sind gegen genetische und umweltinduzierte Variationen und Störungen abgepuffert: Man spricht von „developmental capacitance“, was mit „Aufnahmefähigkeit“ oder „Belastbarkeit“ der Entwicklung übersetzt werden könnte. Wir sprechen im Folgenden von „Entwicklungs-Belastungsfähigkeit“. Gemeint ist, dass die ontogenetische Entwicklung ein gewisses Maß an Störungen von außen (Umwelt) und von Fehlern im Erbgut durch Korrektursysteme ausgleichen und damit abpuffern und verkraften kann. Diese an sich störenden Variationen im Erbgut sind damit vor Selektion geschützt, weil ihre Auswirkungen sozusagen ausgefiltert werden und nicht in Erscheinung treten. Sie machen sich also nicht im Phänotyp bemerkbar. Dieser Effekt wird als Kanalisation der ontogenetischen Entwicklung bezeichnet, weil die Entwicklung trotz gewisser Störungen in die richtige Richtung verläuft. Soweit nun diese Puffersysteme genetische Änderungen abschirmen, können solche Änderungen sich im Erbgut ansammeln. Dadurch kann neue kryptische (verborgene) Variation hinzukommen. Wenn nun aber bestimmte Schwellen der verkraftbaren Umweltstörungen überschritten werden, also die Fehlerbelastung zu hoch wird, kann die kryptische Variation phänotypisch „durchbrechen“ und in Erscheinung treten, womit sie auch der Selektion ausgesetzt wird (die am Phänotyp ansetzt). Sollte der „offengelegte“ Phänotyp adaptiv sein, wird er selektiv gefördert, und es kann schließlich zu Unabhängigkeit von der Umweltänderung kommen (genetische Assimilation). Bevor dieses Konzept weiter erläutert wird, soll dazu ein Beispiel gebracht werden.
Das Beispiel des Hitzeschockproteins Hsp90. Hsp90 ist ein Chaperon, das heißt es sorgt für korrekte Faltung von Proteinen; viele dieser Proteine haben Aufgaben in der Signalübertragung wichtiger Entwicklungswege. Es puffert gegen umweltbedingte Fehler ab, z. B. wenn ein temperaturabhängiger Fehler in der Proteinfaltung auftritt. Ebenfalls sorgt es dafür, dass trotz genetischer Variation die Faltung korrekt erfolgt (stiller Polymorphismus*). Entsprechend sorgt der Ausfall von Hsp90 für viele Schäden, da die Fehler und Störungen nicht mehr korrigiert werden. Dadurch wird die kryptische genetische Variation „offenbar“.
Mit solchen Vorgängen verbinden manche Forscher nun die Hoffnung, dass das Freiwerden kryptischer Variation in bestimmen Umwelten (insbesondere bei Umweltstress) gelegentlich auch einmal vorteilhaft und damit adaptiv sein könnte. Zugleich ist damit eine schnelle Antwort auf veränderte Selektionsbedingungen möglich. Nach einiger Zeit können Änderungen unabhängig von der Mutation von Hsp90 werden (genetische Assimilation). Solange Hsp90 intakt ist, kann sich kryptische Variation ansammeln (deren Wirkung durch Hsp90 ausgeschaltet wird und die dadurch nicht der Selektion ausgesetzt ist). Rutherford & Lindquist (1998, 336) sehen in diesem Vorgang einen plausiblen Mechanismus, „evolutionäre Veränderungen in sonst fest verwurzelten Entwicklungsprozessen voranzubringen“. Sie halten es sogar für möglich, dass durch das plötzliche Freiwerden kryptischer genetischer Variation „die schnellen morphologischen Radiationen, die im Fossilbericht überliefert sind, ermöglicht werden“ (S. 341). (Hier wird einmal mehr deutlich, dass das plötzliche Erscheinen von ganzen Formengruppen in der Fossilüberlieferung als evolutionär problematisch betrachtet wird; andernfalls müsste man ja nicht nach besonderen Erklärungen suchen.) Ähnlich vertreten Cowen & Lindquist (2005, 2185) die Auffassung, dass Hsp90 eine Rolle in der Evolution dadurch spielen könnte, dass es als ein Puffer wirkt, um genetische Variation zu sammeln und freizugeben. Abgesehen von der Frage, wie realistisch dieses Szenario überhaupt ist (s. u.) gibt es einen dicken Wermutstropfen: Homozygote Mutationen von Hsp90 sind letal (tödlich). Hsp90-Mutationen könnten sich folglich nur bis zu einem bestimmten Prozentsatz in der Population durchsetzen. Diese Situation erinnert an die Situation bei der Sichelzellenanämie, bei der homozygote Mutationen letal, heterozygote in malariabelasteten Gegenden dagegen vorteilhaft sind. Anders als dort ist ein Vorteil von Hsp90-Mutanten im heterozygoten Zustand bislang allerdings rein hypothetisch.
|
|
„Developmental capacitance“ und die Entstehung evolutiver Neuheiten
Kanalisation und Pufferung gegen Fehler werden also aus evolutionärer Perspektive als wichtig erachtet, weil diese Mechanismen das Ansammeln genetischer Variation ermöglichen, die zunächst vor der Selektion geschützt wird, später aber freiwerden und vielleicht adaptiv sein kann. Nun kommt es darauf an, dass durch eine Störung der Puffersysteme kryptische Änderungen phänotypisch sichtbar werden und zugleich zufällig adaptiv und dadurch selektierbar sind. Und gleichzeitig (oder kurz danach) muss eine Mutation eintreten, die diese Änderung fixiert (genetische Assimilation). Auf diese Weise sollen neue Phänotypen evolutiv formbar sein.
„Entwicklungs-Belastungsfähigkeit ermöglicht daher den Ursprung, die Akkumulation und schließlich die Freigabe kryptischer genetischer Variation als Antwort auf genetische und Entwicklungsstörungen“ (Moczek 2008, 442). Dieses hypothetische Evolutions-Szenario beschreibt Masel (2005) wie folgt: „Diese Mechanismen haben das Potential, Evolvierbarkeit zu fördern, in dem Sinne, dass potenziell adaptive phänotypische Variation enthüllt wird zu einer Zeit, in der sie am meisten gebraucht wird. Sie agieren als „Kondensatoren“. Folgen wir dieser Analogie, dann wird Variation, die allmählich durch Mutation erzeugt wird, in versteckter Form durch den Kondensator gesammelt. Wenn sie benötigt wird, kann diese Variation freigegeben werden, und die gelegentliche Freigabe verborgener Variation könnte zu evolutionären Innovationen führen“ (Hervorhebung nicht im Original). Damit wird die Latte sehr hoch gelegt, wenn auf diese Weise die Entstehung evolutionärer Neuheiten erklärt werden soll.
|
1. Ein verstecktes Modifikationsspektrum könnte Rohmaterial für nachfolgende Evolution beinhalten. -> Führung phänotypischer Veränderungen vor genetischen Änderungen. 2. Eine neue Modifikationsmöglichkeit wird durch eine stressende Umwelt zur Ausprägung gebracht (Änderung des Phänotyps). 3. Die neue Modifikation muss nachfolgend genetisch fixiert werden: Erhöhung der Umweltsensitivität: kryptische genetische Variation muss herausgeholt werden (Änderung des Genotyps). 4. Durch nachfolgende genetische Assimilation wird die neue modifikative Änderung durch eine Mutation genetisch dauerhaft fixiert. Was kann eine stressende Umwelt aus einem Organismus herausholen – phänotypisch und genetisch?
Ein Beweis für Evolution durch genetische Akkomodation erfordert nach Moczek (2007, 303) den Nachweis der vier folgenden Schritte: 1. Eine Population war nicht in der Lage, bestimmte Phänotypen zu evolvieren, bis 2. Umweltstörungen einen solchen Phänotyp (oder wenigstens einen Vorläufer) auslösten, welcher 3. wiederum durch das Freiwerden kryptischer genetischer Variation begleitet wurde, 4. die selektiert wurde, was schließlich diesem Phänotyp erlaubte, über Generationen hinweg vererbbar zu sein.
|
Vorteil Geschwindigkeit? Wie oben bereits bei „Genetische Akkommodation“ erwähnt, wird in diesem Vorgang gegenüber der klassischen Sicht ein großer Vorteil darin gesehen, dass die Änderungen sich viel schneller durchsetzen können. Denn dem Umweltreiz, der kryptische genetische Variation herausholt, sind viele Individuen auf einmal ausgesetzt; daher besteht eine viel schnellere Ausbreitungsmöglichkeit in der Population, als wenn Mutationen sich allmählich durchsetzen müssen. „Die plötzliche und reversible Freigabe eines Variationsbereichs ist fundamental verschieden von der allmählichen Einführung von Variation durch Mutation“ (Masel 2005, 1359); siehe aber den dritten Punkt in der nachfolgenden Kritik.
Kritik. Nicht nur auf den ersten Blick wirkt das beschriebene Szenario sehr unplausibel. So stellt Moczek (2007, 301) selbst fest, dass so wie die Mehrheit der Mutationen neutral oder schädlich ist auch die große Mehrzahl der stressinduzierten neuen Phänotypen vermutlich nicht-adaptiv ist. Das ist noch freundlich formuliert, denn es ist doch sehr fraglich, dass bei Ansammlung selektionsneutraler Änderungen Positives entstehen sollte. Und die kryptische Variation, die durch Puffersysteme vor der Selektion geschützt ist, ist zudem nur solange neutral, wie sie verborgen ist; sonst wäre sie höchstwahrscheinlich schädlich. Die Schäden beim Ausfall von Hsp90 sprechen diesbezüglich eine klare Sprache. Wie sollen also durch mehrheitlich schädliche oder bestenfalls neutrale Änderungen Umprogrammierungen der Entwicklungswege erfolgen, so dass sogar evolutionäre Innovationen auftreten, wie Masel (2005) mutmaßt (s. o.)? Moczek (2008, 441) meint, versteckte genetische Variation sei selektiv neutral und frei, sich anzusammeln. Das gilt aber nur, solange sie vor Selektion geschützt wird. Es gibt keinen empirischen Grund zur Annahme, dass die angesammelte Variation dann plötzlich konstruktiv sein sollte, wenn sie durch einen Auslöser infolge von Umweltstress freigesetzt wird.
 |
| Wie sollen durch mehrheitlich schädliche oder neutrale Änderungen Umprogrammierungen der Entwick-lungswege erfolgen, so dass sogar evolutionäre Innovationen auftreten? |
 |
|
Man müsste, wie Moczek (2008, 443) schreibt, auf das Zusammentreffen zweier glücklicher Zufälle hoffen: 1. Durch Zufall ändert eine bestimmte Umweltstörung die Entwicklung so, dass sie einen angepassten Phänotyp produziert. 2. Durch Zufall wird kryptische genetische Variation freigegeben, deren Selektion den neuerlichen adaptiven Phänotyp stabilisieren könnte. Dann könnte Evolution durch genetische Akkommodation geschehen und umweltinduzierte phänotypische Variation vererbbar machen (Moczek 2008, 443, „durch Zufall“ ist auch im Original hervorgehoben). Schon der erste Zufall ist extrem unglaubhaft. Moczek (2008, 443) schreibt selbst: „Eine offene Frage bleibt, wie oft das wirklich in der Natur vorkommt und ob es an sich in der Lage ist, die Sorte von bedeutenden neuen Eigenschaften hervorzubringen, deren Ursprung wir zu erklären versuchen.“ Das Problem besteht, wie Masel (2005, 1359) bemerkt, darin, dass alle Kosten eines Ansammelns kryptischer, potentiell innovativer Variation sofort getragen werden müssen, während die (nur eventuell gegebenen) Vorteile in der Zukunft liegen. Zukunftsorientierung aber gibt es in der Evolution nicht.
Es kommt noch ein drittes Problem hinzu: Die genetische Fixierung passender genetischer Akkomodationen erfordert entsprechende Mutationen und diese müssen sich in der Population durchsetzen. Der genannte Geschwindigkeitsvorteil ist damit auf lange Sicht nicht ersichtlich.
Vorprogrammiertes epigenetisches Variationspotential? Vielleicht kann man aber in einer anderen Richtung plausibler spekulieren: Möglicherweise gibt es vorprogrammierte verborgene Polymorphismen, die durch epigenetische Steuerung (z. B. durch stressende Umwelt ausgelöst) sichtbar gemacht werden können. Dass es kryptisches Variationspotential gibt, ist nicht Teil dieser Spekulation; das ist nachgewiesen. Dass es aber vorprogrammiert sein könnte, konstruktive Reaktionen auf Umweltstress zu ermöglichen, erscheint dagegen gewagt. Doch damit wäre nicht das Zusammentreffen äußerst unwahrscheinlicher Zufälle erforderlich, sondern „nur“ eine echte Vorprogrammierung. Dies erfordert eine teleologische Perspektive (Zielorientierung), womit der Rahmen des Evolutionsparadigmas verlassen werden muss.
Dank: Judith Fehrer verdanke ich einige wertvolle Hinweise.
|
Literatur
- Baldwin JM (1896)
- A New Factor in Evolution. Amer. Nat. 30, 441-451, 536-553.
- Cowen LE & Lindquist S (2005)
- Hsp90 potentiates the rapid evolution of new traits: drug resistance in diverse fungi. Science 309, 2185-2189.
- Junker R (2008)
- Evo-Devo: Schlüssel für Makroevolution? Teil 1: Ausgangspunkt und Anerkennung eines ungelösten Evolutionsproblems. Stud. Int. J. 15, 69-75.
- Junker R (2009)
- Evo-Devo: Schlüssel für Makroevolution? Teil 2: Wiederverwendung, Umfunktionierung und Neuprogrammierung. Stud. Int. J. 16, 17-21.
- Masel J (2005)
- Evolutionary capacitance may be favored by natural selection. Genetics 170, 1359-1371.
- Moczek AP (2007)
- Developmental capacitance, genetic accommodation, and adaptive evolution. Evol. Dev. 9, 299-305.
- Moczek AP (2008)
- On the origins of novelty in development and evolution. BioEssays 30, 432-447.
- Myers PZ (2006)
- Evolution of a polyphenism. http://scienceblogs.com/pharyngula/2006/02/evolution_of_a_polyphenism.php (Zugriff 27. 3. 2009).
- Nanjundiah V (2003)
- Phenotypic plasticity and evolution by genetic assimilation. In: Müller GB & Newman SA (eds) Origination of Organismal Form. Beyond the Gene in Developmental and Evolutionary Biology (Vienna Series in Theoretical Biology). Cambridge, MA, pp 245-262.
- Pennisi E (2006)
- Hidden genetic variation yields caterpillar of a different color. Science 311, 591.
- Pigliucci M, Murren CJ & Schlichting CD (2006)
- Phenotypic plasticity and evolution by genetic assimilation. J. Exp. Biol. 209, 2362-2367.
- Rutherford SL & Lindquist S (1998)
- Hsp90 as a capacitor for morphological evolution. Nature 396, 336-342.
- Stern DL & Emlen DJ (1999)
- The developmental basis for allometry in insects. Development 126, 1091-1101.
- Suzuki Y & Nijhout HF (2006)
- Evolution of a polymorphism by genetic accomodation. Science 311, 650-652.
- Waddington CH (1953)
- Genetic assimilation of an acquired character. Evolution 7, 118-126.
- West-Eberhard MJ (2005a)
- Phenotypic accommodation: adaptive innovation due to developmental plasticity. J. Exp. Zool. 304B, 610-618.
|
|
|  |
