 |
Überraschender Befund: Gras gab es schon
zur Zeit der Dinosaurier
Schöpfungstheoretische Aspekte zum frühen Auftreten von Gräsern von Manfred Stephan
Studium Integrale Journal
13. Jahrgang / Heft 1 - Mai 2006
Seite 22 - 30
|
 |
Zusammenfassung: Nachdem bisher makroskopische Fossilien von Gräsern (Poaceae) erst aus dem Bereich der Paläozän/Eozän-Grenze (Alttertiär) bekannt waren, beschreiben nun Prasad et al. (2005) Grasreste bereits aus der höchsten Oberkreide (obere Maastrichtium-Stufe) von Indien. Damit ist gleichzeitig erstmals belegt, daß auch vierbeinige Dinosaurier (Titanosaurier) neben anderen Pflanzen Gräser verzehrten, denn die Reste wurden in ihrem fossilen Kot entdeckt. Die Gräser umfassen mehrere Arten und gehören beiden Obergruppen der Gräser an. Im evolutionstheoretischen Szenario wird deshalb der Ursprung der Gräser noch weiter zurückverlegt, nämlich in die mittlere Oberkreide (Santonium-/Campanium-Stufen).
Im Gegensatz zu den reichen Blätter- bzw. Baumfloren im gesamten Tertiär sind Gräser noch bis zum Mittelmiozän (Jungtertiär) sehr selten und werden erst ab Obermiozän (allmählich) bestandsbildend; dazu erfolgt ein Überblick über mitteleuropäische Fossillagerstätten. Säugetiere mit typischen hochkronigen (hypsodonten) Grasfressergebissen treten vermehrt erst ab Mittelmiozän auf; jedoch gibt es bereits in der Oberkreide und im Alttertiär (Eozän und Oligozän)
einige Gruppen von Säugetieren mit hochkronigem Gebiß.
Abschließend wird das sporadische Auftreten (fehlende bis geringe Bestandsbildung) von Gräsern im Alttertiär und erst recht in der Oberkreide im Rahmen der biblisch-urgeschichtlichen Geologie mit dem Konzept der „geologisch nicht überlieferten Biotope“ gedeutet. Darunter sind Lebensräume zu verstehen, deren Überreste in der Abfolge der Schichtgesteine nicht auftauchen. Deshalb fehlen auch Fossilien der Lebewesen, die in diesen Biotopen existierten. In diesem Zusammenhang werden hilfreiche Aspekte von Fossil-Effekten aus der Paläontologie angesprochen (z.B. der Fossil-Trap-Effekt).
| |
|
Einleitung
Gräser (Poaceae oder Gramineae) sind heute mit über 10.000 lebenden Arten so alltägliche Gewächse, daß man sie kaum von der Erdoberfläche wegdenken kann. Jeder kennt den vielfach teppichartig gepflegten Monokultur-Rasen unserer Schreber- und Vorgärten. Für viele Landschaften Mitteleuropas bestimmend sind Wiesen, Weiden und Hochgebirgs-Almen. Nicht zu vergessen der die Arktis umspannende Tundra-Gürtel, die ausgedehnten afrikanischen Savannen und asiatischen Steppen, die südamerikanischen Pampas oder die nordamerikanischen Prärien (soweit sie nicht in Anbaugebiete verwandelt wurden; vgl. Stanley 1994, 555). Beinahe kann man sagen: Was auf dem Festland nicht Trocken- oder Kältewüste ist, nicht Wälder, Ackerland, Wohngebiete oder Verkehrswege, ist bestandsbildend von Gräsern besiedelt. Und doch handelt es sich überraschenderweise bei Gräsern um Pflanzen, die – geologisch-paläontologisch gesehen – bestandsbildend erst in ausgesprochen späten Sedimenten auftreten, und zwar beginnend mit dem Obermiozän (Jungtertiär; radiometrisch datiert seit höchstens 10 Ma; vgl. Tab. 1). Im Alttertiär (Eozän und Oligozän) sind Gräser sehr selten; das gilt auch noch bis zum Mittelmiozän (bis vor ~ 11 Ma; Piperno & Sues 2005; s.u.). Zwar kennt man vermutliche Graspollen (Monoporites) aus der obersten Stufe der Oberkreide (Maastrichtium) und der untersten Stufe des Tertiärs (Paläozän) Südamerikas, Indiens und Nordafrikas, aber unzweideutige Makrofossilien der Gräser erscheinen nicht früher als im Oberen Paläozän (mit ca. 55 Ma angegeben; Collinson et al. 1993, 830; Prasad et al. 2005, 1177; vgl. Stanley 1994, 522/523). Demgegenüber sind die an Schilf (Phragmites) erinnernden Gräser, die Mägdefrau (1968, 354) reichlich bereits aus dem Teufelsmauer-Sandstein (mittlere Oberkreide) bei Blankenburg (nördl. Harzvorland) beschreibt, sozusagen „Ausreißer“. Sie gehören jedoch nicht zu den flächendeckenden Gräsern; häufiger werden sie aus dem Tertiär beschrieben (s.u.).
 |
| Tab. 1: Links: In der Abfolge der geologischen Systeme sind die Systeme hervorgehoben, in denen Gräser auftreten (Obere Oberkreide, Tertiär und Quartär). Rechts: Die grasführenden Systeme sind in Stufen untergliedert (Maastrichtium, Paläozän etc.). Die Zahlen in Klammern (abgerundet) markieren das derzeit angenommene Alter des Beginns der Stufen in million annus (lat. Jahr). So liegt die Untergrenze des Paläozän und zugleich die Kreide/Tertiär-Grenze bei ~ 65 Ma. Hervorgehoben sind die wichtigsten Schritte des vermuteten und tatsächlichen Auftretens der Gräser. Ferner ist das Erscheinen von Säugetieren mit hochkronigen Zähnen notiert, die an Grasnahrung angepaßt sind. Gräser treten unter den Bedecktsamern (Angiospermen) vergleichsweise spät auf (Obere Oberkreide), während Blätter- bzw. Baumfloren der Angiospermen schon seit der Unterkreide bekannt sind (dazu Kutzelnigg 2000, 52-56). Teilweise ist vermerkt, wenn Gräser weithin oder komplett in geologisch unbekannten Biotopen existierten (Oberkreide und Alttertiär). Besonders auffallend ist die späte Flächenbesiedlung (Savannenbildung) der Gräser: erst ab Obermiozän. |
|
|
Gräserfunde in der jüngsten Oberkreide – Dinosaurier fraßen auch Gras
Nun hat ein indisch-schwedisches Forscherteam eindeutige Reste von Gräsern mit typischen Kieselsäure-Einlagerungen in der obersten Oberkreide (oberes Maastrichtium) entdeckt (Prasad et al. 2005), und zwar in fossilem Saurierkot (s.u.). Nach Piperno & Sues (2005) belegt diese Arbeit erstmals unzweideutig, daß sich die Gräser schon während der Kreidezeit ausgebreitet haben. Die Funde stammen aus Fluß- bzw. See-Sedimenten (fluviatil-lakustrin) der Lameta-Formation bei Pisdura in Zentral-Indien (radiometrisch mit 65-67 Ma datiert). Diese am Fundort etliche Meter mächtige Schichtenfolge ist den ausgedehnten Basalt-Decken des Deccan-Vulkanismus zwischengeschaltet (vgl. Jäger 1998, 185/186). Es handelt sich um mikroskopisch kleine Teile gut erhaltener, Kieselsäure-imprägnierter Grasreste einschließlich Gewebe und Zellen. Die fossilisierende Kieselsäure-Imprägnierung ist von primären, zu Lebzeiten erfolgten Kieselsäure-Einlagerungen (Phytolithe) zu unterscheiden. Letztere sind für viele Gräser kennzeichnend und auch in zwei der gefundenen Gräser enthalten. Im indischen Nand-Dongargaon-Bezirk führt die Lameta-Formation Graspollen.
 |
Der Ursprung der Gräser wird in
die mittlere Oberkreide zurück verlegt;
dort sind sie aber bisher
nicht fossil überliefert. |
 |
|
Interessant ist, daß die zentralindischen Grasreste aus fossilem Kot (Koprolithen) gewonnen wurden, und daß der Kot offenbar von vierfüßigen Echsenbecken-Dinosauriern (Saurischia; Sauropoda) stammt, die zur Gruppe der Titanosaurier gehören (vgl. Müller 1985, 431f.). Ihre Skelettreste sind seit langem aus der Lameta-Formation bekannt; sie gehören zu den letzten Dinosauriergruppen der höchsten Oberkreide und lebten hier nicht lange vor ihrem (wohl) endgültigen Aussterben an der Kreide/Tertiär-Grenze (vgl. Pálfy 2005, 142-152). Die unerwartete Entdeckung, daß bereits Dinosaurier Gras verzehrten, war dem renommierten Wissenschaftsjournal Science neben dem Hauptartikel (Prasad et al. 2005) auch einen Kommentar wert (Piperno & Sues 2005).
Der fossile Kot enthält nicht nur Gräser, sondern auch versteinerte Pflanzenhaare zweikeimblättriger Bedecktsamer (dicotyledonte Angiospermen) sowie „typisch mesozoische“ Pflanzenelemente wie Reste von Nadelbäumen (Coniferen) und Palmfarnen (Cycadeen); dazu kommen Bakterienkolonien, Pilzsporen und Algen. Daraus wird auf eine sehr gemischte Pflanzennahrung der titanosauriden Dinosaurier geschlossen. Ihre Zähne waren auch nicht hochkronig (hypsodont) und dementsprechend nicht als eigentliche Grasfressergebisse ausgebildet.
Nach Piperno & Sues (2005, 1127) ist bemerkenswert, daß bei den Funden zumeist Gräser fehlen, die als ursprünglich bzw. primitiv (basal) interpretiert werden (dazu gehören Gattungen wie Anomochloa, Streptochaeta und Pharus), während Angehörige typischer heutiger Formen (Puelioideae, Ehrhartoideae und Bambusoideae) häufig sind. Taxonomisch werden Gräser (Poaceae) in zwei Obergruppen unterteilt, die BEB- und die PACCAD-Gruppe. Da in der Lameta-Formation (höchste Oberkreide) von Zentral-Indien bereits Arten aus beiden Großgruppen gefunden wurden, schließen Prasad et al. (2005) sowie Piperno & Sues (2005) aufgrund der neuen Daten im Denkrahmen der Makroevolution auf eine weiter zurückliegende rasche Aufspaltung (Radiation) der großen Gruppen der Gräser, die vor mehr als 80 Ma (mittlere Oberkreide; etwa Santonium/Campanium) stattgefunden habe (vgl. Tab. 1).
|
|
Unter frühen Säugetieren gab es ebenfalls Grasfresser
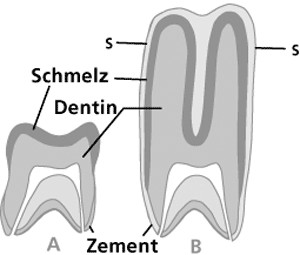 |
| Abb. 1: Niedrigkroniger Zahn (A) und hochkroniger (hypsodonter) Zahn (B) mit säulenförmig verlängerten Höckern, tief eingefaltetem Zahnschmelz und vollständigem Zementüberzug. Im Gegensatz zum niedrigkronigen bleibt beim hochkronigen Backenzahn das Kauvermögen durch Zahnschmelz und Zement voll erhalten, wenn er (etwa bis zur Linie s-s) durch die harte Kieselsäureeinlagerung der Gräser mehr und mehr abgekaut wird. Das gilt erst recht, wenn der Zahn lange Zeit oder sogar zeitlebens nachwächst. (Nach Romer & Parsons 1991). |
|
Piperno & Sues (2005, 1128) nehmen an, daß titanosauride Dinosaurier nicht die einzigen Grasfresser während der Oberkreide waren, sondern auch bestimmte frühe Säugetiere dieser Zeit dazu in der Lage gewesen sind. Besonders die rätselhaften Säugetiere von Gondwana, dem einstigen Großkontinent der Südhalbkugel, die über hochkronige Backenzähne verfügten, kämen dafür in Frage. Denn weil Gräser harte Kieselsäure in ihr Gewebe einbauen, werden die Zähne der Grasfresser stark abgenutzt. Das typische Grasfressergebiß von Säugetieren besteht daher aus hochkronigen, mit Schmelzleisten verstärkten Zähnen, deren Wurzeln sich spät oder überhaupt nicht schließen (Abb. 1). Sie wachsen lange Zeit oder lebenslänglich nach, um die starke Abnutzung der Zähne auszugleichen (Müller 1989, 29-32; Romer & Parsons 1991, 315-317).
Nicht nur in der Oberkreide, auch im Alttertiär (Eozän und Oligozän) sind Grasfunde noch selten (s.u.). Dennoch besitzen gewisse alttertiäre Säugetiere Gebisse, die offenbar auf Grasnahrung eingestellt sind. So sind bestimmte Nashornartige (Amynodontiden) aus dem Obereozän und Oligozän mit „mäßig“ bis „deutlich“ hypsodonten Backenzähnen ausgestattet (Müller 1989, 535f.). Nach Thenius (1979b, 188) werden sie „meist als Sumpfbewohner gedeutet, doch dürften es eher Savannen- und Steppenformen gewesen sein“, also in Grasarealen gelebt haben. Bestimmte großwüchsige Sternhuftiere (Astrapotheria; vgl. Thenius 1979a, 471), die vom Obereozän bis zum Miozän in Südamerika auftreten, verfügen zum Teil über hochkronige Backenzähne (Thenius 1979b, 219). Noch ausgeprägtere zeitliche „Ausreißer“ mit hochkronigen Zähnen sind die Stylinodontinen Nordamerikas, die mit dieser Zahnform schon im Untereozän auftreten. Sie gehören zur Huftiergruppe der Taeniodonta und stehen isoliert im System der Säugetiere, weshalb sie von Thenius (1979b, 241) im Anhang behandelt werden. Ihr hochspezialisiertes Gebiß aus hypsodonten Vorder- und Backenzähnen ist wurzellos sowie bandförmig mit Schmelz bedeckt; deshalb werden sie auch Bandzähner genannt (Müller 1989, 605-609).
|
|
Mitteleuropäische Beispiele: Gräser – selten vom Alttertiär bis zum Mittelmiozän
Das seltene Vorkommen von Gräsern wird schon an einigen mitteleuropäischen Fundorten deutlich. So sind aus berühmten Fossillagerstätten wie dem Geiseltal bei Halle (Saale) oder der Grube Messel bei Darmstadt (beide Mitteleozän; Alttertiär; radiometrisch auf ca. 49 Ma datiert; Harms et al. 2001, 158) anhand von Pollen und Blättern eine große Anzahl Baumarten beschrieben worden, die zu den Blütenpflanzen (Bedecktsamer; Angiospermen) gehören (Überblick z.B. bei Krumbiegel et al. 1982, 77-90; Schaarschmidt 1988, 31-52). Aber es werden kaum Funde von Gräsern gemeldet. Ihre „Pollen treten im Alttertiär äußerst selten auf“; sie kommen „z.B. in Messel noch nicht vor. Im Geiseltal treffen wir Gräserpollen erst ganz vereinzelt“ (Schaarschmidt 1988, 32; vgl. Krumbiegel et al. 1982, 90). Daran haben auch neuere Grabungen in Messel kaum etwas geändert (z.B. Harms et al. 2001, 164); gleiches gilt für das im letzten Jahrzehnt verstärkt erforsche, wenig jünger eingestufte, aber ebenfalls mitteleozäne Eckfelder Maar in der Westeifel (z.B. Lorenz & Lutz 2004, 163; vgl. Boy et al. 2005, 230).
Auch im Oligozän ändert sich dieser Befund kaum. Aus dem obersten Oligozän von Rott bei Bonn (mit ca. 25 Ma datiert), dessen Pflanzenfunde „sehr umfangreich“ sind (v. Koenigswald 1994, 150), führt Mägdefrau (1968, 403) nur Grasfunde an, die an die Gattung Panicum erinnern (vgl. Piperno & Sues 2005, 1127). Auch die umfangreiche Datensammlung pflanzenführender Fossillagerstätten aus ganz Mitteleuropa von Hantke (1993) gibt für das Oberoligozän von Rott nur Insekten an, die an Gräsern leben (S. 191); ferner werden aus dem Oligozän von Mulhouse (Ostfrankreich) Gräser erwähnt (S. 126). Feuchtigkeitsliebende Schilf- (Phragmites) und Ried-Gräser nennt Hantke etwas häufiger; jedoch gehören sie nicht zu den flächendeckenden Gräsern.
 |
| Abb. 2: Modell des unter- bis obermiozänen „Waldpferdes“ Anchitherium. Sein niedrigkroniges Gebiß war nur für Blattnahrung, nicht für harte Gräser geeignet. Schulterhöhe 1,30 m. (Aus Heizmann et al. 1996; Abdruck mit freundlicher Genehmigung). |
|
Aus dem Untermiozän des Randecker Maares (am Rand der Schwäbischen Alb bei Kirchheim/Teck; mit 16-17 Ma angegeben), einer „der reichsten jungtertiären Pflanzenfundstellen in Baden-Württemberg, ja in Deutschland überhaupt“, das über 100 Arten aus 40 Pflanzenfamilien geliefert hat, erwähnen Schweigert et al. (1998, 30, 52) keine Gräser, wohl aber Hantke (1993, 132). Hier wurde das eselgroße dreizehige pferdeartige Anchitherium gefunden (Abb. 2). Man nimmt an, daß es sich bei einer untermiozänen (auf ca. 18 Ma datierten) Säugetier-Wanderungsbewegung von Amerika über die Bering-Landbrücke (zwischen Alaska und Ostsibiren) nach Asien und weiter nach Europa ausgebreitet hat (Heizmann et al. 1996, 9; Steininger 2000, 171). Die niedrigkronigen Zähne weisen auf einen Laubäser hin (Schweigert et al. 1998, 51).
Die ins mittlere Mittelmiozän eingestufte Sedimentfüllung des Steinheimer Beckens (östliche Schwäbische Alb; mit 13-14 Ma angegeben) führt eine reiche Blätterflora. In diesem Becken (das der Einschlag eines kosmischen Körpers hinterließ) bildete sich ein See; für ihn wird nach den Funden ein „dichter Riedgürtel aus Schilf (Phragmites) und Riedgräsern (Cladiocarya)“ rekonstruiert (Heizmann & Reiff 1998, 171; vgl. Heizmann 2002, 152, 98). Nach Gregor (1983, 9) fanden sich Reste von grasähnlichen Blättern überall in den Sedimenten von Steinheim. Erwähnt werden „parallelnervige ‚schilfartige’ Blattreste“, die er zu den Riedgräsern (Cyperaceae; s.o.) stellt, sowie weitere unsichere grasartige Formen. Auch hier tritt das blattäsende „Waldpferd“ Anchitherium auf (Heizmann 2002, 126).
Alle süddeutschen Fundstellen werden jedoch in ihrem Pflanzenreichtum von Öhningen (bei Stein am Hochrhein) übertroffen (Mittelmiozän; mit etwa 13 Ma angegeben); etwa „500 Pflanzenarten sind aus den feingeschichteten Wechsellagen von Kalken und Mergeln“ überliefert (Heizmann & Schmidt 1998, 186). Nach Mägdefrau (1968, 419) liegen in Öhningen von Gräsern „viele Abdrücke vor; besonders häufig war ein Schilfrohr (Phragmites)“. Wie im Steinheimer Becken besteht also auch hier eine weitgehende Fehlanzeige bei flächendeckenden Gräsern.
Nach der oben erwähnten, umfangreichen Datensammlung von Hantke (1993) werden die Funde flächendeckender Gräser erst im Mittel-/ Obermiozän häufiger: Senftenberg/Lausitz (S. 61); Wackersdorf/Oberpfalz (S. 257); Innviertel/Österreich (S. 291); West-Karpaten (S. 314); Burgenland/Österreich (S. 323); Steiermark (S. 332/334). Ähnlich ist es im jüngsten Tertiär (Pliozän): Thüringen (S. 63); Willershausen am Harzrand (S. 89); Allershausen im Solling (S. 91); Westkarpaten (S. 315). Weit häufiger sind die Angaben über quartäre Grasfunde; sie werden hier nicht aufgeführt.
Natürlich geben solche Sammlungen nicht alle Daten vollständig wieder; ferner sind erfahrungsgemäß auch unzuverlässige stratigraphische Einstufungen, Bestimmungsfehler, Verwechslungen usw. nicht auszuschließen. Dennoch scheint insgesamt das Gesamtbild erstaunlich konsistent zu sein: Erst ab Obermiozän/Pliozän ist mit einer stärkeren, zunehmend flächendeckenden Verbreitung der Gräser zu rechnen.
|
|
Vermehrtes Auftreten von Säugetier-Grasfressern ab Obermiozän
 |
| Abb. 3: Skelett des obermiozän-pliozänen „Urpferdes“ Hippotherium. Als typischer Grasfresser war es mit hochkronigen Zähnen ausgestattet, die denen heutiger Pferde stark ähneln. (© Staatliches Museum für Naturkunde Stuttgart) |
|
Eine stärkere Änderung der Säugetierfauna ist in den ältesten Ur-Rhein-Sanden von Eppelsheim in Rheinhessen (frühes Obermiozän; auf etwa 11 Ma datiert) durch das vermehrte Auftreten von Grasfressern unter den Säugetieren zu verzeichnen (Tabelle mit Huftieren z.B. bei Rutte 1987, 39; vgl. Rothausen & Sonne 1984, 61-63). Neben dem oben erwähnten Anchitherium tritt hier das ebenfalls dreizehige pferdeähnliche Hippotherium auf (Abb. 3); es wurde früher Hipparium genannt (Boy 2005, 217; v. Koenigswald 1994, 175; vgl. 1998, 152 Anm. 9). Beide überlappen sich nur während einer (kurzen) Zeit im Obermiozän Europas, nach v. Koenigswald (1998, 143) zwischen 11,5 bis 8,5 Ma. Man geht davon aus, daß Hippotherium sich bei einer späteren Einwanderungswelle (mit etwa 11,5 Ma angegeben) von Amerika aus über die Beringbrücke nach Asien und Europa ausgebreitet hat (Steininger 2000, 171), und zwar „sehr schnell“, denn „die in Europa auftretende Art ist fast unverändert gegenüber der amerikanischen Ausgangsform“ (v. Koenigswald 1998, 143). Hippotherium unterscheidet sich von den unter- bis obermiozänen „Waldpferden“ der Gattung Anchitherium durch seine „hochkronigen, an Grasnahrung angepaßten Zähne“; sie ähneln „sehr stark“ denen heutiger Pferde (Heizmann 1992, 59). Der Einschnitt in die Säugetierwelt, der mit dem plötzlichen Auftreten von Hippotherium in Europa verknüpft ist, wird nach dem alten Gattungsnamen „Hipparion event“ genannt (Heizmann et al. 2003, 6).
Auch in Spaltenfüllungen bei Salmendingen und Melchingen (Schwäbische Alb) werden Anchitherium und Hipparion zusammen gefunden. Allerdings ist in diesem speziellen Fall (senkrechte Spaltenfüllungen!) die völlig gleichzeitige Einschwemmung von Skelettresten in offene Spalten der Weißjura-Oberfläche nicht gesichert (vgl. zum Problem solcher Faunenvermischungen Stephan 2002, 199).
Wie in Eppelsheim (s.o.) ist bei den wenig jünger datierten Sedimenten am nördlichsten Hegau-Vulkan, dem Höwenegg bei Immendingen, eine stärkere Änderung der Säugetier-Fauna zu verzeichnen (frühes Obermiozän; mit 10 Ma angegeben; Heizmann et al. 2003, 6). Die Flora wurde nach Schweigert (1998, 205) noch nicht systematisch untersucht; er erwähnt aus „feuchten Bereichen“ auch hier „Riedgräser, wie das Schneidried (Cladium)“ bzw. „Schilfachsen“ (Heizmann et al. 2003, 12). Diese bedeutende Fundstelle lieferte besonders zahlreiche Reste des aus Eppelsheim bekannten, pferdeartigen dreizehigen Hippotherium; bis zum Ende der damaligen Ausgrabungen (1958) wurden 13 vollständige Exemplare geborgen (Rutte 1987, 50; vgl. Schweigert 1998, 204). Neben Hippotherium gehören zu den neu eingewanderten Faunenelementen u.a. grasfressende Antilopen der Gattung Miotragocerus (Heizmann 1992, 59). Reste davon wurden in der Pilotgrabung (2003) der zurzeit laufenden neuen Grabungskampagne etwa gleich häufig wie Hippotherium gefunden (Heizmann et al. 2003, 13). Im Gegensatz zu den Verhältnissen, wie sie noch zur Mittelmiozän-Zeit herrschten, muß man sich nach Schweigert (1998, 205) „die Vegetation im Obermiozän nun deutlich stärker aufgelockert vorstellen … wodurch auch ein Lebensraum für die in Herden lebenden Großsäugetiere, wie grasfressende Pferde und Antilopen, geschaffen war“ (vgl. v. Koenigswald 1998, 189f.). Allerdings scheint bei solchen Rekonstruktionen Vorsicht geboten zu sein. Nach v. Koenigswald (1994, 175) besitzt eine kleine obermiozäne Blattflora, die bei Sprendlingen (zwischen Frankfurt/M. und Darmstadt) gefunden wurde, „keinerlei Hinweise auf ein Trockenklima, in dem sich eine Savanne bilden konnte“. Dieser Befund könnte ein Hinweis darauf sein, daß sich auch im Obermiozän die Savannen erst (sehr) begrenzt ausgebreitet hatten.
Noch jünger als die o.g. Schichten von Eppelsheim, ins höchste Obermiozän, werden die isolierten Urrhein-Sedimente von Dorn-Dürkheim zwischen Alzey und Oppenheim eingestuft (mit 8,7 bis 5,4 Ma angegeben; Franzen 1999, 201; vgl. Boy et al. 2005, 219). Es sind Ablagerungen, die nach Rutte (1987, 53) aus „einer genetisch unerklärlichen Mischung von Sanden, Tonen, Geröllen“ aufgearbeiteter älterer Sedimente bestehen, mit „massenhaften (zerbrochenen) Skelettresten überwiegend sehr großer Säugtiere“. Auch die geologische Position des kleinräumigen Vorkommens ist „außergewöhnlich und schwer erklärbar“, da man vor seiner Bildung lokal große tektonische Mobilität (starke Hebung) annehmen muß (Rothausen & Sonne 1984, 64/65). Den gräserdominierten Lebensraum, der aus dem Bau der gefundenen Säugtiere abgeleitet wird, kommentiert Rutte (1987, 53) knapp so: „Weitgehend alle Bewohner einer Savannenlandschaft“.
|
|
Zusammenfassender Überblick: Ökologischer Wechsel im Miozän
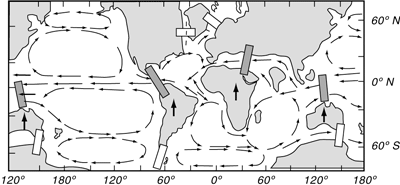 |
| Abb. 4: Vereinfachte Modellvorstellung der Änderung von Meeresströmungen im Mittel/ Jungtertiär als vermutete Teilursache einer komplexen, mehrstufig verlaufenden globalen Abkühlung: Durch die unterschiedlich schnelle Nordwanderung der Südkontinente Australien, Afrika, Indien und Südamerika (mit Pfeilen nach Norden) und ihre Kollision mit Eurasien (und Nordamerika) wurde die tropisch-subtropische Ost-West-Warmwasserzirkulation der Ozeane nach und nach unterbrochen (blaue Balken). Die gleichzeitige Öffnung der Meereswege zwischen der Antarktis und den nordwärts driftenden Südkontinenten (helle Balken) isolierte die Antarktis vom subtropischen Meerwasser. Zunehmend wurde der Kontinent von einem kalten Strömungsring umgeben; seit dem Miozän bildete sich Inlandeis. Über Meeresströmungen erhielten die weitgehend voneinander isolierten Ozeane (Atlantik, Indik, Pazifik) mehr und mehr kühles Oberflächenwasser vom antarktischen Strömungsring. Zudem füllten sich die Tiefen der Ozeanbecken komplett mit absinkendem antarktischem Kaltwasser. (Nach Wefer & Berger 2001). |
|
Nach Stanley (1989, 199) gehört die Ausbreitung von Grasflächen auf Kosten der Blattfloren zu den „auffallendsten Florenwechseln“. Sie wird u.a. auf eine Klimaverschlechterung (Abkühlung) zurückgeführt, ein sehr komplexes Geschehen, für das man mehrere Ursachen in zeitlich gestaffelten Schüben annimmt, die hier nur teilweise und umrißhaft genannt werden können. Als Auslöser gelten zum einen die Nordwanderung der Südkontinente (Afrika, Arabien und Indien) und die dadurch bedingte Änderung von Meeresströmungen infolge Schließung der ozeanischen Ost-West-Verbindung (Tethys-Ozean – Mittelmeer) beim Andocken der Südkontinente an Eurasien (Abb. 4). Ferner die durch den Zusammenstoß der Kontinente verursachte, beginnende Heraushebung der alpinen Kettengebirge Eurasiens, vor allem des Himalaya-Tibet-Plateaus. Als weiterer Grund werden wiederholte Meeresspiegelschwankungen und die Ausbreitung der Gletscher auf der Antarktis mit der dadurch bedingten Zunahme kalten Tiefenwassers in den Ozeanbecken genannt (z.B. Steininger 2000, 167-171; Buggisch & Walliser 2001, 46-48; Wefer & Berger 2001, 87-96).
Für die obermiozäne Ausbreitung von Kräutern und Gräsern wird folgendes generelles paläobiologisches Szenario angenommen: Die Verbreitung erfolgte in etwa gemeinsam mit der Ausbreitung der Pferdeartigen und vieler Huftiere, die hochkronige Backenzähne besaßen und damit besonders gut feste Gräser mit ihren harten Kieselsäure-Einlagerungen verwerten konnten (s.o.). Zu den Huftieren zählen Rinder, Antilopen, Schafe und ihre Verwandten (Bovidae). Auch unter den miozänen Nashörnern sind „manche auf Grasnahrung spezialisiert“; dazu gehört ebenso ihre Hochbeinigkeit, die für rasche Fortbewegung in offenen Landschaften vorteilhaft ist. „Da grasreiche, offene Parklandschaften erst im Obermiozän in Europa eine stärkere Rolle spielen, sind derart angepaßte Arten im Untermiozän noch selten“ (Heizmann et al. 1996, 42, 44). Bestimmte Paarhufer und Schlieferartige (Hyracoidea), die ab Mittelmiozän und im Pliozän verbreitet waren, erreichten mindestens Tapirgröße; sie hatten hypsodonte bzw. „partiell hypsodonte“ Backenzähne (Thenius 1979b, 211, 222). In Nordamerika ist die (mikroevolutive) Radiation und artenreiche Ausbreitung von grasfressenden Pferdeartigen ab dem Mittelmiozän sehr ausgeprägt (vgl. z.B. Gould 1998, 87, 90-94). Insekten, die an die neuausgebreiteten Gräser und Kräuter angepaßt waren, verbreiteten sich, aber auch Nagetiere und viele Singvögel, die sich von Samen und Insekten ernähren. Das Gleiche gilt für Giftschlangen, die wiederum Nagetiere wie Mäuse und Ratten sowie Vogeleier in großer Zahl fressen (Stanley 1989, 200f; Stanley 1994, 555f; vgl. Heizmann 1992, 60; Heizmann et al. 1996, 10).
|
|
Gräser – zeitweise in unbekannten Lebensräumen
Die beschriebenen Befunde können so zusammengefaßt werden: Säugetiere mit typischem, hochkronigen Grasfressergebiß treten vermehrt ab Mittelmiozän auf. Bedeutsam ist am fossilen Hervortreten der Gräser: Großflächige Bestandsbildung („Savannen“) entstand offenbar erst im Obermiozän/Pliozän; demgegenüber waren Gräser im Eozän und Oligozän äußerst selten. Bisher kannte man die ältesten unzweideutigen Makro-Fossilien von Gräsern ungefähr von der Paläozän-Eozän-Grenze, aber vermutliche Graspollen schon aus der obersten Oberkreide (Maastrichtium) und dem untersten Alttertiär (Paläozän). Dazu kommt nun der gesicherte Fund von Blattresten mehrerer Grasarten in der höchsten Oberkreide (Ober-Maastrichtium), und zwar von beiden Großgruppen der Gräser. Aus dieser bereits prätertiären Vielgestaltigkeit (Piperno & Sues 2005, 1127) schließen Prasad et al. (2005) im Rahmen des evolutionären Szenarios auf einen gemeinsamen Vorfahren der Gräser in der mittleren Oberkreide (Santonium/ Campanium; mit mehr als 80 Ma angegeben).
Im mittleren und jüngeren Alttertiär (Eozän und Oliozän) waren also Gräser nur sehr untergeordnete Bestandteile der Pflanzenwelt, verglichen mit den überaus reichen Funden an Blatt- bzw. Baumfloren. Dennoch gibt es aus diesen Schichtfolgen Säugetiergruppen mit hochkronigem (hypsodonten) Grasfressergebiß. Säugetiere mit diesem Zahnbau sind sogar schon aus der Oberkreide bekannt (Piperno & Sues 2005, 1128). Dieser Befund dürfte zumindest regionale grasbestandene Flächen im Alttertiär voraussetzen, die jedoch geologisch-paläontologisch so gut wie nicht (direkt) überliefert sind (s.u.). Wie die neuen Gräserfunde belegen, gilt das auch für die mittlere Oberkreide. Denn für diese Zeit (Santonium/Campanium) postulieren Prasad et al. (2005) aus evolutionstheoretischen Gründen das Vorkommen verschiedener Gräser (s.o.).
Insgesamt ist der Mangel an Grasfunden und besonders Pollen im Alttertiär und der späten Oberkreide, erst recht aber ihr Fehlen in der mittleren Oberkreide erstaunlich. Pollen sind zwar klein, aber sehr widerstandsfähig (Mägdefrau 1968, 31). Denn die äußere Zellwand von Sporen und Pollen „wird zu großen Teilen aus Sporopollenin, das sind hochpolymere Ester von Fettsäuren und Carotinoiden, aufgebaut. Sporopollenin gilt als eine der widerstandsfähigsten bekannten organischen Substanzen“ (Heunisch 1999, 207). Wenn z.B. tertiäre Pollen durch Umlagerungsvorgänge in quartäre Sedimente gelangen (sog. Zombie-Effekt; vgl. Schatz 2000), sind sie dennoch gut identifizierbar (vgl. Hantke 1993, 17). Sie werden zudem jährlich in großen Mengen produziert und mit dem Wind oft weit verbreitet. Pollenanalysen sind in der Literatur in großer Zahl publiziert. Auch wenn Pollen von Gräsern etwa in Fossillagerstätten des Tertiärs mit ihren reichen Blatt- bzw. Baumfloren (z.B. Messel oder Geiseltal) nicht unbedingt reichlich erwartet werden, sollte man annehmen, daß sie von weither eingeweht wurden – es sei denn, sie waren zu der Zeit außerordentlich selten.
|
|
Geologisch nicht überlieferte Biotope als Erklärungsansatz – Paradebeispiel Dinosaurier
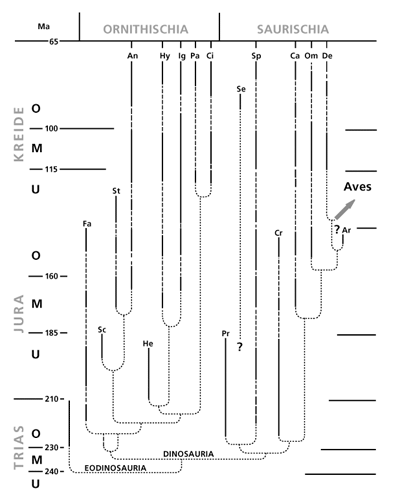 |
| Abb. 5: Fossilüberlieferung der Dinosaurier (Dinosauria). Sie werden in der Evolutionslehre auf Reptilien zurückgeführt, die als Eodinosauria zusammengefaßt sind (unten). Diese hätten sich in die beiden Hauptgruppen, Vogelbecken-Dinosaurier (Ornithischia) und Echsenbecken-Dinosaurier (Saurischia), aufgespalten (oben). Der hypothetische Stammbusch an der Basis ist jedoch nicht fossil belegt (angedeutet durch punktierte Linien). Die gestrichelten Teile der Balken markieren ebenfalls erhebliche Lücken in der Fossilüberlieferung. Sie treten mehrfach in der Abfolge auf, und zwar bei allen weit durchlaufenden Dinosaurierlinien. Links außen: Geologische Systeme, in deren Schichtfolgen die Dinosaurier auftreten (Trias usw.). Großbuchstaben daneben markieren den unteren, mittleren und oberen Abschnitt der Systeme. Daneben die derzeitig angenommenen Alter in Millionen Jahren (Ma; oben). Rechts: Pfeil deutet die vermutete evolutive Abzweigung der Vögel (Aves) an. (Nach Haubold 1990) |
|
Wenn die fossilen Überreste von Lebewesen (-Gruppen) in bestimmten Schichtgliedern nicht gefunden werden, obwohl sie in älteren und jüngeren Schichtfolgen auftreten, erklärt das auch die evolutionsorientierte Paläontologie mit geologisch nicht überlieferten Lebensräumen, also fossil unbekannten Biotopen, in denen diese Organismen lebten: „Wichtiger ist aber noch, daß es Zufluchtsorte gegeben haben muß, …von denen wir nichts wissen“ (Pálfy 2005, 191). Noch wichtiger ist die Konsequenz daraus: „Das bedeutet, daß längst nicht alle zu einer bestimmten Zeit existierenden Biotope auch fossil überliefert werden“ (Dietl & Schweigert 1999, 46; Überblick bei Stephan 2002, 61-121; 128-130). Das gilt erstaunlicherweise sogar für einen Teil der Dinosaurier-Biotope in den sehr gut untersuchten, nicht-marinen (terrestrischen) Schichtfolgen von Obertrias, Jura und Kreide, wie Haubold (1990, 208; vgl. 32-34) ermittelt hat: „Während des Mesozoikums ist der Anteil der Lücken, wie bei nahezu jeder Fossilgruppe, erheblich und bleibt eine stets zu beachtende Komponente bei verallgemeinernden Interpretationen“ (Hervorhebung nicht im Original; Abb. 5). Nicht nur bei den Dinosauriern, praktisch in der gesamten Fossilüberlieferung ist ein Paradox zu beobachten: Obwohl sich durch fortschreitende Forschung immer wieder kleinere Lücken schließen, bleibt insgesamt der Befund großer Lückenhaftigkeit bestehen. Denn weit vor dem bisher bekannten Erscheinen bestimmter Lebewesengruppen werden des öfteren überraschend gleichartige „Vorläufer“ entdeckt (Dietl & Schweigert 1999, 40; Stephan 2002, 128/129), und ebenso erscheinen plötzlich lange nach dem bisherigen Verschwinden aus der Fossilüberlieferung („Aussterben“) Angehörige der gleichen Gruppe als „Nachläufer“ (z.B. Schweigert & Dietl 1999, 9; Dietl & Schweigert 2001, 45). Damit sind zwischen den Neufunden und den schon bekannten gleichartigen Fossilien wieder neue Lücken entstanden. Offenbar hat die Lückenhaftigkeit systematischen Charakter, denn sie ist praktisch in jeder Fossilgruppe zu beobachten. Bei den Dinosauriern kommt hinzu: Obgleich längst nicht alle riesig waren, äußerte sich ihre Dominanz „in einer alle Lebensbereiche der Kontinente beeinflussenden, kontrollierenden und gestaltenden Funktion“ (Haubold 1990, 13). Deshalb ist es besonders bedeutsam, daß sogar ihre Fossilüberlieferung erhebliche Lücken aufweist. Auch diese Befunde „zeigen uns, wie unvollständig im Grunde die Fossilüberlieferung der einstigen Lebewelt ist“ (Dietl & Schweigert 2001, 52).
|
„Das bedeutet, daß längst nicht alle
zu einer bestimmten Zeit existierenden
Biotope auch fossil überliefert werden.“ |
|
|
Doch zurück zu den Gräsern. Bisher wurden die ältesten Funde in der obersten Oberkreide entdeckt; jedoch treten hier gleichzeitig mehrere Arten aus beiden Obergruppen der Gräser auf (s.o.). Deshalb schließt auch die evolutionstheoretisch orientierte Paläontologie auf noch weiter zurückliegende, aber unbekannte Lebensräume von Gräsern (weitere Beispiele solcher Rückschlüsse: Stephan 2002, 122-127; 131-133). Als Ursprungszeit der Gräser nimmt man die mittlere Oberkreide (Santonium/Campanium) an. Bestätigend kommt in diesem Fall hinzu, daß aus der Oberkreide Säugetiere mit hochkronigem Gebiß beschrieben worden sind (s.o.); es ist daher plausibel, für sie auf Grasnahrung zu schließen. Demnach kann auch von Gräsern in geologisch unbekannten Lebensräumen der mittleren Oberkreide gesagt werden, „daß solche mindestens als Seltenheiten und damit unauffindbar stets existierten“ (Jaeger 1991, 316).
|
|
Paläontologische Effekte: Zum Verhalten von Fossilgruppen
In neuerer Zeit werden bestimmte Effekte des Fossilbefundes diskutiert, die solche Phänomene, zumindest teilweise, erklären könnten. Paläontologen begründen öfters das anfängliche fossile Fehlen einer neu erscheinenden Lebewesengruppe damit, daß ihre ältesten Mitglieder viel zu selten sind. In den Schichtfolgen, die zu Beginn ihres Erscheinens abgelagert wurden, ist deshalb das Auftreten bzw. Auffinden solcher Fossilien statistisch außerordentlich unwahrscheinlich; das hat man als „Spill-Rongis-Effekt“ bezeichnet. Dieser Effekt steht für das umgekehrte Phänomen (und wird deshalb rückwärts gelesen!) wie der häufiger in der Literatur auftauchende Signor-Lipps-Effekt, der nach seinen Erstbeschreibern, zwei Paläontologen, benannt wurde (Pálfy 2005, 187-189, 26-28). Letzterer erklärt die abgestufte Reihenfolge des Verschwindens verschiedener Organismen bei einem Aussterbeereignis: Die (sehr) seltenen Arten verschwinden als erste aus der Fossilüberlieferung. Die (etwas) stärker verbreiteten Formen finden sich noch höher in der Schichtenfolge. Nur die dominierenden, zahlreichen Fossilien treten (fast) bis ans Ende des (angenommenen) endgültigen Aussterbe-Einschnitts auf. Unzureichende Fossildokumentation infolge mangelhafter Sammlungsmethodik durch die Bearbeiter kann ebenfalls einen (viel) zu frühen Aussterbe-Zeitpunkt vortäuschen (Gould 2002, 198-206).
Der „Spill-Rongis-Effekt“ bezieht sich auf Probleme beim fossilen Erscheinen von Lebewesen, und der Signor-Lipps-Effekt auf ihr Verschwinden (Aussterben). Um letzteres geht es auch beim Rarity-Effekt: Bei fossilen Arten, deren Überreste zwar geographisch weit verbreitet sind, aber generell sehr selten gefunden werden, ist der Zeitpunkt des Aussterbens noch schwerer zu bestimmen. Beim Fossil-Trap-Effekt betrifft die Unsicherheit nicht nur die Aussterbezeit, sondern die Gesamtexistenzdauer einer Art: Denn hier geht es um Arten, die nur sporadisch in sehr kleinräumigen Fossillagerstätten gefunden werden, also lediglich punktuell bekannt sind (Schatz 2000). Solche Organismen haben „ein ganz bestimmtes, nur selten vorkommendes oder fossil überliefertes Habitat besessen“ (Schweigert 2002, 8), d.h. sie besiedelten einen besonderen, kleinräumigen Lebensraum. Diese Biotope besitzen daher „nur ein sehr geringes geologisches Überlieferungspotential“ (Schweigert & Dietl 1999, 9).
Da Gräser zeitweilig wegen ihrer großen Seltenheit nicht fossil erscheinen, ist – analog zu den genannten Fossil-Effekten – eine Aussage über ihre Existenzdauer vor ihrem fossilen Auftreten ebenfalls kaum möglich. Die anhand der Gräser beschriebenen Befunde passen gut in das Konzept der „geologisch nicht überlieferten Lebensräume“ (Stephan 2002, 61-147), das im vorigen und in diesem Abschnitt beschrieben wird. Dieses Konzept wird hier im Rahmen der biblisch-urgeschichtlichen Geologie angewendet, d.h. es wird versucht, es im Kurzzeit-Horizont der biblischen Urgeschichte zu verstehen (Stephan & Fritzsche 2003, 157-187). Dabei wird vorgeschlagen: Geologisch nicht überlieferte Lebensräume, die z.B. anhand der genannten Fossileffekte angesprochen wurden, sollten nicht nur für (einzelne) geologische Stufen, sondern überhaupt für die geologischen Systeme bedacht werden. Und zwar unter folgender Annahme: Die betreffenden Organismen bildeten während längerer Zeit nur kleine Populationen, die zudem eine paläogeographisch sehr geringe Ausdehnung hatten. Statistisch ist es außerordentlich unwahrscheinlich, daß solche Lebensräume fossile Reste hinterlassen haben bzw. daß diese jemals entdeckt werden. Auch im Deutungsrahmen der Evolutionslehre sieht sich die Paläontologie aufgrund solcher Befunde häufig genötigt, die längere fossile „Abwesenheit“ von Organismen, die mehrere geologische Systeme umfassen kann, nicht nur bei einzelnen Fossilarten, sondern zum Teil auch bei umfangreichen Fossilgruppen anzunehmen (z.B. Binder & Stephan 2005; Junker 2002; Junker & Stephan 2004; Stephan 2002; 2005, 86).
|
Literatur
- Binder H & Stephan M (2005)
- Zwischen gigantischen Tausendfüßern und kleinen Pinselfüßern: Fossile Lücke von 200 Millionen Jahren. Stud. Int. J. 12, 39-40.
- Boy J et al. (2005)
- Geologie von Rheinland-Pfalz. Hg Landesamt für Geologie und Bergbau; Mainz. Stuttgart.
- Buggisch W & Walliser OH (2001)
- Erdgeschichte als Klimageschichte. In: Huch M, Warnecke G & Germann K (Hg) Klimazeugnisse der Erdgeschichte – Perspektiven für die Zukunft. Berlin, S. 17-49.
- Collinson ME, Boulter MC & Holmes PL (1993)
- Magnoliophyta (‚Angiospermae’). In: Benton MJ (Ed) The Fossil Record 2; London, S. 809-841.
- Dietl G & Schweigert G (1999)
- Nusplinger Plattenkalk. Eine tropische Lagune der Jura-Zeit. Stuttgarter Beitr. Naturk. C 45, 1-62.
- Dietl G & Schweigert G (2001)
- Im Reich der Meerengel. Der Nusplinger Plattenkalk und seine Fossilien. München.
- Franzen JL (1999)
- Die große Flut – der Rheinhessensee. Natur und Museum 129, 201-212.
- Gould SJ (1998)
- Illusion Fortschritt. Die vielfältigen Wege der Evolution. Frankfurt/M.
- Gould SJ (2002)
- Ein Dinosaurier im Heuhaufen. Streifzüge durch die Naturgeschichte. Frankfurt/M, S. 193-209.
- Gregor HJ (1983)
- Die miozäne Blatt- und Frucht-Flora von Steinheim am Albuch (Schwäbische Alb). Dokumenta naturae 10; München, 1-45.
- Hantke R (1993)
- Flußgeschichte Mitteleuropas. Skizzen zu einer Erd-, Vegetations- und Klimageschichte der letzten 40 Millionen Jahre. Stuttgart.
- Harms F-J, Aderhold G, Hoffmann I, Molek H, Nix T & Rosenberg F (2001)
- Grube Messel bei Darmstadt (Südhessen). Jber. Mitt. oberrhein. Geol. Ver., N.F. 83, 145-183.
- Haubold H (1990)
- Die Dinosaurier. System, Evolution, Paläobiologie. Die neue Brehm-Bücherei 432. Wittenberg Lutherstadt.
- Heizmann EPJ (1992)
- Das Tertiär in Südwestdeutschland. Stuttgarter Beitr. Naturk. C 33, 1-61.
- Heizmann EPJ (2002)
- Leben nach der Katastrophe – Paläontologie eines Meteorkraters. In: Heizmann EPJ & Reiff W: Der Steinheimer Meteorkrater. München, S. 81-159.
- Heizmann EPJ, Duranthon F & Tassy P (1996)
- Miozäne Großsäugtiere. Stuttgarter Beitr. Naturk. C 39, 1-60.
- Heizmann EPJ, Munk W, Ziems A, Bernor RL & König H (2003)
- Neue Grabungen am Höwenegg (Gemeinde Immendingen, Landkreis Tuttlingen, Baden-Württemberg). Carolinea 61, 5-16.
- Heizmann EPJ & Reiff W (1998)
- Aus der Katastrophe geboren – Das Steinheimer Becken. In: Heizmann EPJ (Hg) Vom Schwarzwald zum Ries. Erdgeschichte mitteleuropäischer Regionen, 2. München, S. 165-176.
- Heizmann EPJ & Schmidt FX (1998)
- Tertiärer Vulkanismus auf der Schwäbischen Alb und im Hegau. In: Heizmann EPJ (Hg) Vom Schwarzwald zum Ries. Erdgeschichte mitteleuropäischer Regionen, 2. München, S. 177-190.
- Heunisch C (1999)
- Die Bedeutung der Palynologie für Biostratigraphie und Fazies in der Germanischen Trias. In: Hauschke N & Wilde V (Hg) Trias – eine ganz andere Welt. Mitteleuropa im frühen Erdmittelalter. München, S. 207-220.
- Jaeger H (1991)
- Neue Standard-Graptolithenzonenfolge nach der „Großen Krise“ an der Wenlock/Ludlow-Grenze (Silur). N. Jb. Geol. Paläont. Abh. 182, 303-354.
- Jäger M (1998)
- Faunenschnitt – Was geschah am Ende der Kreidezeit? (Teil 2) Fossilien 15, 181-189.
- Junker R (2002)
- Erste Landpflanzen und molekulare Uhren. Stud. Int. J. 9, 42-43.
- Junker R & Stephan M (2004)
- Die ältesten Salamander sind „modern“. Stud. Int. J. 11, 31-32.
- Koenigswald Wv (1994)
- Obermiozäne Säugetiere aus dem Ur-Rhein von Eppelsheim. In: Koenigswald Wv & Meyer W (Hg) Erdgeschichte im Rheinland. Erdgeschichte mitteleuropäischer Regionen 1. München, S. 173-178.
- Koenigswald Wv (1998)
- Säugetierreste aus Karsthohlräumen der Schwäbischen Alb. In: Heizmann EPJ (Hg) Vom Schwarzwald zum Ries. Erdgeschichte mitteleuropäischer Regionen 2. München, S. 139-152.
- Krumbiegel G, Rüffle L & Haubold H (1982)
- Das eozäne Geiseltal – Ein mitteleuropäisches Braunkohlenvorkommen und seine Pflanzen- und Tierwelt. Die neue Brehm-Bücherei 237. Wittenberg Lutherstadt.
- Kutzelnigg H (2000)
- Das „abscheuliche Geheimnis“ – Woher kommen die Angiospermen (Teil 1). Stud. Int. J. 7, 51-58.
- Lorenz V & Lutz H (2004)
- Das quartäre Meerfelder Maar, das eozäne Eckfelder Maar bei Manderscheid und die eozänenen Flussablagerungen von Gut Heeg in der Westeifel. Jber. Mitt. oberrhein. Geol. Ver., N.F. 86, 125-185.
- Mägdefrau K (1968)
- Paläobiologie der Pflanzen. 4. Aufl. Jena.
- Müller AH (1985)
- Lehrbuch der Paläozoologie, III Vertebraten, Teil 2 Reptilien und Vögel. 2. Aufl. Jena.
- Müller AH (1989)
- Lehrbuch der Paläozoologie, III Vertebraten, Teil 3 Mammalia. 2. Aufl. Jena.
- Pálfy J (2005)
- Katastrophen der Erdgeschichte – globales Artensterben? Stuttgart.
- Piperno D & Sues H-D (2005)
- Dinosaurs Dined on Gras. Science 310, 1126-1128.
- Prasad V, Strömberg CAE, Alimohammadin H & Sahni A (2005)
- Dinosaur Coprolithes and the Early Evolution of Grasses and Grazers. Science 310, 1177-1180.
- Romer AS & Parsons TS (1991)
- Vergleichende Anatomie der Wirbeltiere. Nachdruck der 5. Aufl. Hamburg-Berlin.
- Rothausen K & Sonne V (1984)
- Mainzer Becken. Sammlg. Geol. Führer 79. Berlin-Stuttgart.
- Rutte E (1987)
- Rhein • Main • Donau. Wie – wann – warum sie wurden. Eine geologische Geschichte. Sigmaringen.
- Schaarschmidt F (1988)
- Der Wald, fossile Pflanzen als Zeugen eines warmen Klimas. In: Schaal S & Ziegler W (Hg) Messel – Ein Schaufenster in die Geschichte der Erde und des Lebens, Senckenberg-Buch 64. 2. Aufl. Frankfurt/M, S. 29-51.
- Schatz W (2000)
- Die taxonomische Unschärfe von Aussterbeereignissen. In: Gradualism vs. Catastrophism. Internet (07.05.01): http://www-geol.unine.ch/sgs/Katabstracts, 3-4.
- Schweigert G (1998)
- Alles schon mal da gewesen – Vegetationsgeschichte Südwestdeutschlands von der Tertiärzeit bis heute. In: Heizmann EPJ (Hg) Vom Schwarzwald zum Ries. Erdgeschichte mitteleuropäischer Regionen 2. München, S. 199-208.
- Schweigert G (2002)
- Zwei neue Garnelen (Decapoda: Dendrobranchiata, Eukyphida) aus oberjurassischen Plattenkalken Süddeutschlands. Stuttgarter Beitr. Naturk. B 323, 1-11.
- Schweigert G, Böttcher R, Heizmann EPJ & Wohnhas W (1998)
- Das Randecker Maar. Ein fossiler Kratersee am Albtrauf. Stuttgarter Beitr. Naturk. C 43, 1-66.
- Schweigert G & Dietl G (1999)
- Neubeschreibung von „Eryon longipes O. Fraas“ (Crustacea, Decapoda, Eryonidea) aus dem Nusplinger Plattenkalk (Ober-Kimmeridgium, Schwäbische Alb). Stuttgarter Beitr. Naturk. B 274, 1-19.
- Stanley SM (1989)
- Krisen der Evolution – Artenaussterben in der Erdgeschichte, Spektrum Bibliothek 18. 2. Aufl. Heidelberg.
- Stanley SM (1994)
- Historische Geologie. Eine Einführung in die Geschichte der Erde und des Lebens. Heidelberg.
- Steininger FF (2000)
- Europa im Känozoikum: Die Tertiär-Periode. In: Meischner D (Hg) Europäische Fossillagerstätten. European Palaeontological Association. Berlin, S. 165-171.
- Stephan M (2002)
- Der Mensch und die geologische Zeittafel. Warum kommen Menschenfossilien nur in den obersten geologischen Schichten vor? Holzgerlingen.
- Stephan M (2005)
- Plattenkalke: Neue Hinweise auf rasche Entstehung und geologisch nicht überlieferte Biotope. Stud. Int. J. 12, 85-87.
- Stephan M & Fritzsche T (2003)
- Sintflut und Geologie. Schritte zu einer biblisch-urgeschichtlichen Geologie. 2. Aufl. Holzgerlingen.
- Thenius E (1979a)
- Stammesgeschichte der Huftiere. In: Grzimek B (Hg) Grzimeks Tierleben 12. München, S. 468-473.
- Thenius E (1979b)
- Die Evolution der Säugetiere. Eine Übersicht über Ergebnisse und Probleme. UTB 865. Stuttgart, New York.
- Wefer G & Berger WH (2001)
- Klima und Ozean. In: Huch M, Warnecke G & Germann K (Hg) Klimazeugnisse der Erdgeschichte – Perspektiven für die Zukunft. Berlin, S. 51-107.
|
|  |
