|
Seit einigen Jahren häufen sich Mitteilungen von Dinosaurierfunden mit „sensationeller Weichteilerhaltung“ (z.B. JI et al. 1998, RUBEN et al. 1997). Üblicherweise findet man von dieser ausgestorbenen Tiergruppe lediglich Überreste von Hartgeweben wie mineralisierte Knochen, Zähne und Eischalen sowie die versteinerten Spuren dieser Tiere: Fußabdrücke und ganz selten die als Negativabdruck überlieferte Struktur der Hautoberfläche.
Nicht sehr bekannt ist, dass auch schon zu Beginn des 20. Jahrhunderts Dinosaurierfunde mit Weichteilerhaltung publiziert worden sind (GILMORE 1914). Dies wurde aber in der Fachwelt nicht besonders beachtet. Es ist sogar nachgewiesen, dass aus Unwissenheit, unsachgemäßer Bearbeitung und aus Zeitgründen frühere Dinosaurierfunde von potentieller Weichteilerhaltung „befreit“ wurden, damit man an die begehrten Knochen schneller herankommen konnte.
| |
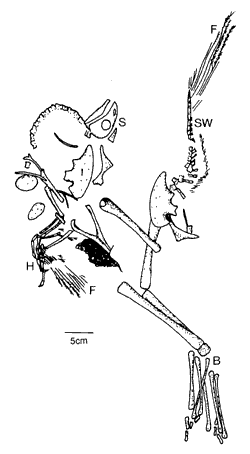 |
| Abb. 1: Skizze des fossilen Theropoden Caudipterxyx. S Schädel, SW Schwanzwirbelsäule, H Handskelett, B Beinskelett, F Federn (Nach JI et al. 1998) | |
Mit dem Bekanntwerden zahlreicher „Dinosaurierfriedhöfe“ in China und der Mongolei seit den 1970er Jahren, nahm auch die Fundmenge von Dinosaurierresten mit sogenannter Hautschatten- und Federerhaltung sowie sonstiger Weichteilüberreste zu. Andere Fundstellen mit außergewöhnlich gut erhaltenen Dinosaurier-Exemplaren kennt man mittlerweile auch aus Italien und den USA. Besonders erwähnenswert sind hierbei die Funde mit Strukturen, die als versteinerte Lungen- oder Herzreste gedeutet wurden (RUBEN et al. 1997). Diese wie auch die sogenannten federähnlichen Hautgebilde waren natürlich besonders interessant, da hier offensichtlich direkte Nachweise für die Warmblüter-Theorie der Dinosaurier und ihre Ahnenschaft zu den Vögeln vorliegen sollten.
Derzeit sind aus sechs Dinosauriergruppen Exemplare mit filamentösen (fädigen) Hautstrukturen beschrieben worden (zusammengefasst in PETERS 2001). Über den genauen Aufbau der filamentösen Strukturen herrscht kein Konsens unter den Paläontologen. Entweder wird von federähnlichen, vergleichbar mit heutigen Borsten- oder Fadenfedern, oder von haarähnlichen Strukturen gesprochen. Je nach theoretischen Hintergrund des untersuchenden Wissenschaftlers sind diese Filamente echte Federn (SCHWEITZER et al. 1999, JI et al. 2001), also Hinweise auf eine direkte Verwandtschaft zu den Vögeln, oder Strukturen, die ein dichtes Haarkleid ähnlich das der Flugsaurier und Säugetiere aufbauen (FEDUCCIA 2001). Besonders interessant ist die Tatsache, dass diese filamentösen Hautgebilde nicht nur bei den gemeinhin als Vogelvorfahren gehandelten Theropoden (fleischfressende Dinosaurier) nachgewiesen worden sind, sondern auch bei den Psittacosauriern vorkommen. Psittacosaurier waren vermutlich rein pflanzenfressende, etwa 2 Meter lange Dinosaurier mit einem papageienähnlichen Schnabel, die in die Nähe der Ceratopsier (Horndinosaurier) gestellt werden. Das Vorkommen der filamentösen Hautstrukturen in diesen unterschiedlichen Gruppen schränkt die Hypothese von der Ahnenschaft der Theropoden aufgrund der nachgewiesenen „Befiederung“ stark ein.
Abbildungen und Beschreibungen dieser Hautgebilde lassen derzeit folgende Schlüsse zu:
- Die Filamente zeichnen sich durch einen überwiegend langgestreckten und unverzweigten „Schaft“ aus.
- Die als leichte Verästelungen beschriebenen Spitzen einiger Exemplare sind auf Grund der Erhaltungsart nicht sicher als primäre Ausbildung zu bestätigen.
- Eindeutige Federtypen wie Kontur- und Deckfedern oder Daunen fehlen bislang oder sind nur bei Gruppen beschrieben worden, die systematisch zu den Vögeln gestellt werden: Archaeopterygidae (z.B. Archaeopteryx), Confuciusornithidae (z.B. Confuciornis), Enantiornithes.
- Die filamentösen Hautgebilde der Dinosaurier ähneln stark den überlieferten Haarstrukturen der ebenfalls ausgestorbenen Flugsaurier.
- Alle Dinosaurier-Arten mit dieser besonderen Hautbedeckung sind geologisch gesehen jünger als der berühmte Urvogel Archaeopteryx aus Bayern. Dadurch scheiden diese als direkte Vorfahren der Vögel aus.
Über die Hautbedeckung der Mehrzahl der nahezu 500 bislang beschriebenen Dinosaurierarten ist nichts bekannt. Von einigen kennt man eine schuppige Hautstruktur ähnlich heutiger Reptilien. Andere kleine und mittelgroße bis ca. 2m lange Dinosaurierarten besaßen vermutlich ein haarähnliches Fell an bestimmten Partien des Körpers und der Extremitäten. Inwieweit diese Hautstrukturen mit den Haaren der Säugetiere identisch sind oder eine ganz eigene Bildung darstellen entzieht sich derzeit unserer Kenntnis. Nach dem jetzigen Kenntnisstand darf aber die Existenz befiederter Dinosaurier angezweifelt werden. Kritisiert werden muss, dass die weitläufig akzeptierte Meinung Vögel als befiederte Dinosaurier zu betrachten, die wissenschaftsneutrale Betrachtung einer erhaltenen Körperbedeckung bereits im Vorfeld genauerer Untersuchungen sehr zugunsten einer Befiederung beeinflusst hat. Nicht selten begegnet man in den Arbeiten der Anhänger der Vogel-Dinosaurier-Abstammungstheorie ein Zirkelschluss der Art, dass basierend auf der Annahme Vögel seien befiederte Dinosaurier, der Nachweis von Federn bei einigen Dinosaurier-Arten diese Grundannahme scheinbar belegt.
|
