Diese bislang etablierte Sichtweise geriet aber zunehmend in Bedrängnis, denn je mehr Daten man anhäufte, um so mehr Widersprüche ergaben sich. Im Zeitalter der modernen Molekularbiologie ist man mittlerweile in der Lage, komplette Genome von Organismen zu vergleichen. Es zeigte sich immer deutlicher, daß sehr viele Ausnahmen von der bisher etablierten Sicht des Evolutionsstammbaumes vorkommen. Es gibt eine erhebliche, nicht zu vernachlässigende Anzahl von Genen, mit denen sich jeweils ein komplett anderer Stammbaum erstellen ließe.
In Kürze gesagt, findet man ein buntes Flickmuster von Genen verschiedenster Herkunft in den einzelnen Organismen. Dieser Befund ist zwar nicht unbedingt neu (vgl. Hilario & Gogarten 1993), wurde aber in seiner Bedeutung bislang unterschätzt. So finden sich z.B. in Archaea Proteine bakterieller Herkunft und eukaryontischer Herkunft. Als Beispiel mag hier das Archaea Methanococcus jannaschii stehen. Etwa 13% der Gene sind ähnlich zu Genen aus Eukaryonten und etwa 44% der Gene ähnlich zu solchen aus Bakterien (Koonin et al. 1997), wie beispielsweise die CoA-Reduktase, Glutamin-Synthetase, das Hitzeschockprotein Hsp70, ATPasen und tRNA-Synthetasen. Diese Enzyme sind nun keineswegs nur in sekundäre Stoffwechselreaktionen eingebunden und somit entbehrlich, sondern erfüllen in diesen Organismen lebensnotwendige, ja zum Teil ganz zentrale Aufgaben.
Unter dem Paradigma einer Schöpfung sind die entstandenen Widersprüche in der Evolutionsgeschichte der Lebewesen erwartet worden (s.o.). Denn ein Designer ist frei in der Verwendung und Anordnung einzelner Komponenten, so lange diese ihre Aufgabe im zellulären Kontext erfüllen können. Dabei ist es unerheblich, ob z.B. eine CoA-Reduktase ein Enzym bakterieller oder eukaryontischer Bauart ist. Diese Sichtweise wurde bereits von ReMine (1993) dargelegt und von diesem als "Design-Signal" gewertet. Man könnte fast meinen, ein Designer sei wie mit einem Salzstreuer über das Organismenreich gegangen.
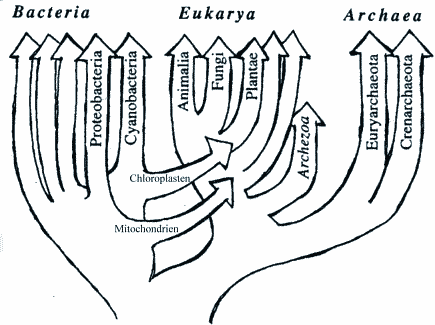
Gespannt durfte man auf die Einordnung der Befunde unter dem Paradigma der Evolutionslehre sein. In einem Übersichtsartikel, erschienen im Juni 1999 im Wissenschaftsmagazin "Science", schlägt Doolittle zur Lösung dieses Problems vor, die Annahme eines universellen gemeinsamen Vorfahren in Form einer primitiven Zelle fallen zu lassen. Er meint statt dessen, daß sich das Leben aus einer komplexen "Urgemeinschaft primitiver Zellen" entwickelt habe, zwischen denen über längere Zeit ein massiver Gentransfer bestand. Der bisher gültige "Stammbaum" verwandelt sich in einem "Stammbusch" mit zahlreichen Überkreuzungen und Vernetzungen (Abb. 2). Dabei treten jedoch zwei schwerwiegende Probleme auf: Wie ereignete sich der Gentransfer und woher kommen die verschiedenen Vorfahren? Massiver Gentransfer zwischen verschiedenen Organismen kann nicht einfach so "passieren". Trotz einer Funktionsverwandtschaft der Enzyme können die Komponenten von komplexen Systemen nicht einfach übertragen werden, sie sind "eingepaßt". Ein Vergleich mag dies illustrieren: Die Kolben eine VW-Käfer-Motors und eines BMW-Motors haben zwar dieselbe Funktion und eine ähnliche Form, aber es dürfte selbst einem geschickten Autobauer schwer fallen, einen VW-Motor mit den Kolben eines BMW-Motors und umgekehrt zu betreiben. Die Ähnlichkeit der Kolben an sich erklärt sich zwanglos durch die Erfindung ("Design") eines ersten Kolbenmotors. Späteren Konstrukteuren stand es dann frei, dieses "Kolbenprinzip" abzuwandeln und ihrem System anzupassen.