|
|
|
|
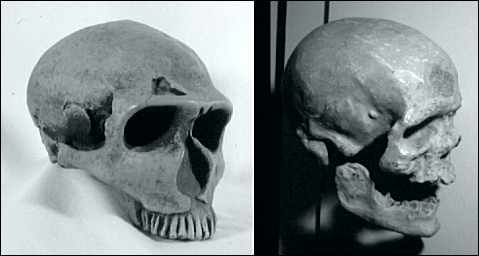
Aufs neue beschäftigt der Neandertaler (Abb. 1) die Gelehrten. In der Paläanthropologie gibt es nach wie vor zwei Strömungen, die sich an der Frage der Artabgrenzung zerstritten haben. Die eine plädiert beim Neandertaler für eine eigene Art, gestärkt unter anderem durch die neuen DNA-Untersuchungen zweier Neandertaler, die eine große Divergenz zum modernen Menschen aufweisen, und damit eine sehr alte eigene Linie nahelegen. Die andere Position nimmt an, daß der Neandertaler nichts weiter als eine merkwürdige Unterart des Homo sapiens auf der evolutiven Reise ins Heute war. Sie wird durch einen möglichen Hybriden zwischen Neandertaler und anatomisch modernen Menschen aus Portugal gestärkt. |
|
Diese Auseinandersetzung hat einen alten Streit zweier Schulen neu angefacht: die Multiregionalisten glauben, daß der anatomisch moderne Mensch zeitgleich und unabhängig voneinander nicht nur in Afrika, sondern auch in Europa, Vorder- und Südost-Asien aus frühen erectus-artigen Homo-Formen entstanden ist (das sogenannte Kandelaber-Modell; der Neandertaler ist ein "Durchgangsstadium" und hat seine Gene in uns hinterlassen). Dagegen glauben die Monozentristen, daß erst relativ unlängst Vorfahren des modernen Menschen Afrika verließen und vorort alle früheren Formen (eben auch den Neandertaler) ohne Kompromisse verdrängten (das Arche-Noah-Modell; der Neandertaler hat keinerlei genetische Spuren in uns hinterlassen). Die Implikationen der neuen Ergebnisse auf die Herkunftsmodelle sind in Tab. 1 zusammengefaßt. |
Als Kommentar zu dieser Veröffentlichung in der renommierten Zeitschrift PNAS erschien Tattersalls engagierte Gegenposition (Tattersall 1999), der eine hitzige Kontroverse folgte. Tattersall hängt der Überzeugung an, daß der Neandertaler eine eigene Art war. Seiner Meinung nach ist der Neandertaler mehr als nur ein Anhängsel und ein uninteressanter Seitenast auf dem Weg zum anatomisch modernen Menschen. Für die allgemeine Beurteilung der menschlichen Evolution sei es nicht unbedeutend, ob man den Neandertaler als eine eigene Art oder nur als eine Subspezies betrachtet. Zudem bestimme eine solche Vorentscheidung die selektive Wahrnehmung nicht unerheblich und beeinflusse wesentlich den Informationsgehalt: Betrachtet man den Neandertaler als eine eigene Art, wird man mehr auf das Besondere achten, umgekehrt würde man sein Augenmerk eher auf das Verbindende richten, hielte man ihn nur für eine Unterart oder gar "Rasse". Da man in der Evolutionsbiologie davon ausgeht, daß nur innerhalb einer Art, d.h. zwischen Unterarten, dauerhaft Hybride gebildet werden können, wäre für Tattersall ein Neandertal-Hybrid schwer zu verkraften. Deshalb geht seine Argumentation dahin, alle beschriebenen Merkmale als für einen anatomisch modernen Menschen dieses Zeithorizontes typisch zu entkräften. Nach Tattersall fehlen alle Hinweise auf einen neandertalartigen Einschlag. |
Der Neandertaler wird als ein europäischer Nachfahre eines erectus-ähnlichen Auswanderers (Homo heidelbergensis) angesehen. Je nach Vorstellung ist er direkte Übergangsform zum anatomisch modernen Menschen oder dessen Seitenzweig mit wenig oder keinem Genfluß (siehe auch Tab. 1). Er bevölkerte zwischen 200.000 und weniger als 30.000 rJ vor unserer Zeit Europa und den Vorderen Orient. Der Neandertaler besitzt tatsächlich eine einzigartige Merkmalskombination, unter Umständen sogar einzelne autapomorphe ("einmalige") Merkmale, die ihn morphologisch von den modernen Menschen abgrenzen. Diese mögen sich schätzungsweise seit ca. 500.000 rJ herausgebildet haben, je nach Hypothesenlage entweder durch eine Neuzuwanderung über den Nahen Osten oder direkt aus den hochvariablen Mittelpleistozänen Mischformen (Homo heidelbergensis), die Europa bevölkerten. Einige Merkmale wie z.B. die enorm hohe Knochenwand-Robustizität, die nur die Armknochen betrifft, stellen die Anthropologen weiterhin vor ein Rätsel (war er ein Schwergewichtler?). Für Tattersall (1999) und viele andere waren deswegen die Ergebnisse der DNA-Vergleiche der vergangenen Jahre keine Überraschung (siehe Kasten). Die großen Unterschiede zwischen Neandertaler-DNA und der des heutigen Menschen werden auf die lange getrennte Geschichte zurückgeführt - und damit auf die Existenz getrennter Arten. Doch die auf den ersten Blick frappierend groß erscheinenden DNA-Unterschiede verlieren angesichts der großen Unterschiede innerhalb anderer Menschenaffen-Arten wieder etwas an Gewicht: die Variabilität, die innerhalb einer einzelner Populationen von wildlebenden Zwergschimpansen festgestellt wurde, ist höher als die der gesamten (!) heutigen Menschheit (Gagneux 1999). Der Bewertungsmodus von morphologischen und molekularbiologischen Unterschieden bleibt weiterhin problematisch. |
Die eigentliche Brisanz des Fundes liegt in dem genannten jahrzehntelangen Streit über den Modus der Evolution zum modernen Menschen (Tab.1). Die Schule der Multiregionalisten sieht die Entstehung des modernen Menschen als einen graduellen Prozeß, der bei der ersten echten Homo-Form begonnen hat und dann gleichzeitig an mehreren Orten der Alten Welt relativ unabhängig voneinander den modernen Menschen, d.h. die jeweils ansässigen Populationen ("Rassen") herausgebildet habe. Diese Hypothese (Multiregional-Modell, Regionale Kontinuität oder auch Kandelaber-Modell genannt) hat wegen potentiell rassistischer Fehlinterpretationen Gegenwind erhalten. Die Befürworter dieser Hypothese, die der synthetischen Evolutionsvorstellung eher nähersteht als die Alternativen, aber von den Daten nicht gut gestützt wird, halten dementsprechend alle Vertreter der Gattung Homo - einschließlich Homo erectus - für monospezifisch: die einzige Art Homo sapiens habe sich zwar als Chronospezies im Evolutionsverlauf verändert, lasse darin aber keine Artabgrenzung erkennen. Irgendwo auf diesem Weg zum modernen Menschen lebte auch der Neandertaler als Glied dieser Kette. Für die Multiregionalisten (Regionale Kontinuität) waren die DNA-Ergebnisse weniger leicht zu verdauen, während der mögliche Hybrid eher bestätigend wirkte. Eine evolutionäre Übergangsform wäre für die Regionale Kontinuität zwar wesentlich interessanter als ein Hybrid, doch würde dieser zumindest als Beleg für die genetischen Nähe der beiden diskutierten Formen dienen. Zuvor hatten schon einige Multiregionalisten verschiedene Fossilien aus dem kritischen Zeitraum zwischen 30.000 und 40.000 rJ als Übergangsformen zwischen Neandertaler und anatomisch modernen Menschen deklariert, doch Tattersall wies alle diese Behauptungen mit Argumenten wie Fehldatierungen bzw. -diagnosen zurück. Die angeführten Kandidaten seien entweder reine Neandertaler oder reine anatomisch moderne sapiens-Formen. Für strenge Monozentristen, die von zwei völlig getrennten Arten mit längerer getrennter Entwicklungsgeschichte ausgehen, ist ein Hybrid eher problematisch, während ihnen die großen genetischen Unterschiede zwischen dem Neandertaler und dem anatomisch modernen Menschen sehr willkommen sind. Die beiden neuen Befunde - große DNA-Unterschiede bei gleichzeitiger Hybridisierungsmöglichkeit - stärken bzw schwächen die zwei ersten Modelle gleichermaßen (Tab. 1), können dagegen von einer Mischform der beiden Modelle als auch vom Grundtypmodell gleichermaßen gut integriert werden. |
Wenn man die Neudatierung der Neandertal-Funde in Kroatien zugrundelegt (Richards 2000), sollte es dort genügend Gelegenheit für Hybridisierungen gegeben haben. Dort lebten noch um 28.000 rJ echte Neandertaler, während schon seit mehreren Tausend rJ anatomisch moderne Menschen durchzogen, die mindestens seit 32.000, wahrscheinlich schon seit 36.000 rJ aus dem Nahen Osten über Zentral-Europa nach Europa einwanderten. Im Zeitraum zwischen 150.000 und 36.000 rJ scheint der Neandertaler in Europa die einzige überlebende archaische sapiens-Form zu sein, bis schließlich nach 40.000 rJ der anatomisch moderne Mensch ("Cro-Magnon"; Abb. 1) aus dem Nahen Osten in sein "Revier" hineindrängte. Nach nur ca. 10.000 rJ war der Neandertaler vollständig verschwunden, in Rückzugsgebieten hielt er sich bis zu 4000 rJ länger (auf der Iberischen Halbinsel kennt man um 27.000 C14-Jahre alte Mousterien-Kulturen aus dem Südspanischen Zafarraya). In den meisten Gebieten Europas läßt sich auch ein entsprechend abrupter Bruch zwischen der Neandertaler Mousterien-Kultur und dem Aurignacien oder anderer Kulturen feststellen, die dem anatomisch modernen Menschen zugeordnet werden. An gewissen Orten jedoch scheinen um 36-33.000 rJ die Neandertaler einige kulturelle und Verhaltensmerkmale der Cro-Magnon-Menschen durch Angleichung (Akkulturation) übernommen zu haben. Ob es tatsächlich eine Akkulturation mit ausführlicher Hybridisierung oder nur ein Berühren zweier Kulturen ohne biologische Folgen war, bleibt offen. Sequenzdaten aus vier Y-Chromosom-Genen heutiger Menschen weisen interessanterweise darauf hin, daß die Populationsgröße des anatomisch modernen Menschen um 28.000 rJ exponentiell zugenommen haben soll: die Schätzung dieses Ereignisses wird mit dem Aussterben/Verschwinden der Neandertaler und der rapiden Ausbreitung des Aurignacien in Verbindung gebracht (Shen 2000). Die verschiedenen Hypothesen, wie und warum es zu einem abrupten Verschwinden des Neandertalers gekommen ist, werden auch zum Teil dadurch geprägt, ob man ihm eine eigene Art zubilligt oder ihn nur als eine Unterart behandelt. Folgende Denkmöglichkeiten werden erörtert: Vom Neandertaler fehlt nach 27.000 rJ jede Spur, weil a. er im kriegerischer Konflikt mit der neuen Menschenform aus dem Nahen Osten unterlegen war, b. er aufgrund geringerer Konkurrenzfähigkeit seine Lebensgrundlage einbüßte, c. er sich durch schnelle Evolution in den anatomisch modernen Menschen verwandelte, d. sein Erbmaterial durch Hybridisierung in das des eindringenden modernen Menschen aufging. Wer sich für zwei Arten stark macht und das Regionale-Kontinuitäts-Modell für falsch ansieht, hält a oder b für wahrscheinlich an. Ein strenger Multiregionalist dagegen hält c für wahrscheinlich, während momentan die meisten Multiregionalisten entsprechend dem Vorschlag von Smith & Spencer (1984) d als am wahrscheinlichsten ansehen. |
Wie auch immer die mosaikartige Natur des Lagar Velho-Kindes in Zukunft gedeutet werden wird und die DNA-Unterschiede bewertet werden, beides ist gut mit den Erwartungen des Grundtypenmodells kompatibel. Wie bei der Evolution bleiben auch hier ähnliche Fragen offen: ist das abrupte Verschwinden der Neandertaler Zeichen von genetischer Absorption, Veränderung oder Verdrängung? Die für den Evolutionsbiologen relevante Frage, in welchem Maße die Neandertaler am Genpool des heutigen Menschen beteiligt waren, muß für die Grundtypenbiologie umformuliert werden: Welchen Grad an genetischer Diversität erreichten Neandertaler und Homo sapiens sapiens im Laufe ihrer Geschichte und welche Folge hatte das z.B. für Fertilität oder Sterilität bei Hybridisierung?
Ein Nachweis für eine Hybridisierung wäre eine Stärkung der Vorstellung, daß alle fossilen Vertreter der Gattung Homo (sensu Wood & Collard 1999) zu einem Grundtyp gehören, wobei das Fehlen desselben gleichermaßen auch als Ausdruck verloren gegangener Hybridisierungsfähigkeit gedeutet werden kann. Ein auch von den Gegenern akzeptierter "Nachweis" von Hybridisierung ist schwer zu führen - es sei denn man findet eine mit mehreren Skelettfunden gesegnete Grabung, die innerhalb eines sehr kurzen Zeitraumes das ganze Kontinuum von reinen Neandertalern, reinen Cro Magnons und der ganzen Schattierung von Hybrid-Merkmalen dazwischen vorweisen kann. Eine schöner Traum für jeden Anthropologen - aber wer weiß? |
Literatur
|

|