|
|
|
|
|
Klassischerweise gibt es zwei Möglichkeiten, die Natur und deren Naturgegenstände (Organismen und ihre Überreste) zu beschreiben: 1. Die naturalistische Einstellung (frühester Vertreter Aristoteles; neuere Literatur: Simpson 1967, Mayr 1975): Man sieht in den Naturgegenständen reale Einheiten, deren Beschreibungen die Natur in Ihrer objektiven Form darlegen. 2. Die handlungstheoretische Einstellung (z.B. Janich 1992, Herkner 1999, Janich & Weingarten 1999): Dem Naturforscher ist sein Eingreifen und die Tatsache, daß er selbst Teil dieses Systems ist, bewußt - Naturgegenstände und Kategorien werden erzeugt. Beide Sichtweisen liefern Ergebnisse, die sich jedoch selbst sehr unterschiedlich definieren. Diese Definitionsfrage ist auch der entscheidende Kritikpunkt. Für den Naturalisten ist der Oberarmknochen (Humerus) bei Wirbeltieren eine tatsächlich in der Natur vorzufindende Struktur, die bei den unterschiedlichen Wirbeltier-Kategorien homologisiert, d.h. wiedergefunden, werden kann. Der handlungstheoretisch operierende Naturforscher ist sich bewußt, daß die Struktur "Humerus" von ihm an einem Individuum ausgewählt und beschrieben worden ist. Der Humerus wäre bei dieser Sichtweise nicht aus sich heraus in der Natur vorhanden, sondern abhängig von der Betrachtung des Beobachters. Noch kniffliger wird es, wenn eine induktive bzw. deduktive Ableitung resultiert oder wenn funktionelle bzw. phylogenetische Aussagen getroffen werden. Im Beispiel bleibend wären so bei naturalistischer Betrachtung alle Naturgegenstände mit einem Humerus näher miteinander verwandt als jene ohne diesen und gehen auf einen gemeinsamen Vorfahren zurück. Diese methodisch nachfolgenden Schritte potenzieren die Grundannahmen. Die kritische Betrachtung, was ein Biologe oder Paläontologe heutzutage tut, wenn er "die Natur" erforscht, ist dringend notwendig, da sich meiner Meinung nach zu viele Naturforscher zu wenig Gedanken über die philosophische Basis ihrer Wissenschaft und ihres Handelns machen. Im Methodenteil von Forschungsarbeiten wird zu diesen Fragen sehr selten Stellung bezogen. Ist man sich automatisch bewußt, nach wessen Sichtweise an die Fragestellung herangegangen worden ist? Was rechtfertigt es, z.B. im Methodenteil einer paläontologisch ausgerichteten Forschungsarbeit nicht zu erwähnen, daß die Grundannahme eine evolutionstheoretisch ausgerichtete ist? Dann hört man hier das Argument, daß die Evolutionstheorie als "bewiesen" gilt und die Erwähnung schlichtweg banal wäre. Ist das wirklich so? Ist Evolution - um bei diesem Beispiel zu bleiben - nicht in der Tat nur ein Gedankengebäude, ein Theorienkomplex, der sicherlich seine Vorzüge an Erklärungsmöglichkeiten liefert, aber nichtsdestotrotz eine Theorie bleibt (im Gegensatz zu Remane 1956). Dies gilt gleichermaßen für evolutionskritische oder kreationistische Aufsätze. Es sollte stets nachvollziehbar sein, nach welchen "Spielregeln" ein Autor vorgegangen ist. Es soll im nachfolgenden aufgezeigt werden, wo die Grenzen der einzelnen Sichtweisen der Naturbeschreibung - insbesondere eines naturalistischen Ansatzes - bestehen. Dabei soll keine Wertung im Sinne von "wahr" oder "falsch" stattfinden, sondern die pragmatischen Seiten herausgestellt werden. |
Die basalen und apikalen Merkmale werden nun als Charakteristika für die entsprechende Kategorie benutzt, wobei apikale Merkmale nur für eine bestimmte Gruppe charakteristisch sein sollen. Aus der Praxis weiß jeder Naturforscher, der eigentlich erst jetzt als Taxonom bezeichnet werden kann (er hat Ordnung geschaffen), daß zwar jede Kategorie durch charakteristische Merkmale definiert ist, welche aber in ihrer Ausprägung keineswegs konstant vorzufinden ist - Stichwort Variabilität. Dieser Begriff beinhaltet letztlich nichts anderes als den Ausdruck einer Vielfalt an Naturgegenständen einer Kategorie. Für die benutzte Methode ergibt sich daraus als logische Konsequenz eine Beschränktheit, diese Vielfalt zu erfassen. Man stelle sich nur einmal vor, ein Naturforscher hätte irgendwann die Möglichkeit, die gesamten Schädel der heute ca. 6 Milliarden lebenden Menschen zu untersuchen. Es wäre zu erwarten, daß der Naturforscher aufgrund der feststellbaren Unterschiede in der Geometrie (Form) und Biometrie (Maße) der Schädel neue, heute noch unbekannte Kategorien aufstellen würde. Statistisch schwankt die Qualität und Quantität eines Merkmals um einen Mittelwert. Je mehr Untersuchungsobjekte einer Kategorie zur Verfügung stehen, desto höher die Wahrscheinlichkeit Variationen in der Merkmalsausprägung festzustellen. Taxonomie kann also folglich niemals ein statisches System sein, sondern unterliegt einer stetigen Veränderung. Ein weiteres Problem besteht darin, daß neben den basalen Merkmalen auch einige vorher als apikale Merkmale betrachtete Charakteristika bei Naturgegenständen unterschiedlichster Kategorien-Zugehörigkeit vorkommen. Diese Erscheinung wird gemeinhin mit Konvergenz umschrieben, die entsprechenden apikalen Merkmale als "konvergent erworbene" Merkmale bezeichnet. Beispielsweise wird die stromlinienartige Körperform von Haien, Delphinen, Ichthyosauriern (Fischechsen) und Pinguinen als ein "konvergentes" Merkmal angesehen. Während die taxonomische Bedeutung bzw. Nichtbedeutung "konvergenter" Merkmale durch andere, scheinbar "echte" apikale Merkmale häufig "überstimmt" wird, sorgt ihr "Dasein" unter Naturforschern, die sich mit Phylogenie beschäftigen, immer wieder für Verwirrung (z.B. Patterson 1982; s. Kap. Taxonomie und Phylogenie). Per Definition haben diese "konvergenten" Merkmale den Status eines basalen Merkmals, da sie in mehr als einer Gruppe vorkommen. Hierbei ist auch folgendes zu beachten: Ein Merkmal kann nur dann als apikal gelten, wenn es in Verbindung mit einer bestimmten Kategorie erwähnt wird. Beispiel: Das Merkmal "Wirbelsäule" ist für die Kategorie "Wirbeltiere" (Vertebrata) ein apikales Merkmal, für die darin eingeschlossenen Unterkategorien "Knochenfische" (Osteichthyes) oder "Vögel" (Aves) jedoch ein basales. Weiterhin folgt aus dem von der Anzahl an untersuchten Naturgegenständen abhängigen System, daß mit dem jetzigen Wissensstand nie mit Sicherheit ein Merkmal als "absolut" apikal angesprochen werden kann. Bis zur Kenntnisnahme aller existierenden und jemals existierten Naturgegenstände bleibt dies nur eine Hypothese. Beispielsweise ist nicht gesagt, ob es nicht doch eine Seegurke oder einen Seeigel mit einer Wirbelsäule gegeben hat oder gibt, was das bestehende Schema kippen würde. Es ist unmöglich und wird auch niemals möglich sein ein, "echtes" apikales Merkmal als solches sicher zu bestimmen. Die logische Konsequenz für ein "konvergentes" Merkmal muß sein, dieses aus der Liste an Charakteristika für die zu definierende Kategorie zu eliminieren. Inwieweit der Taxonom dieses Merkmal zur Aufstellung einer neuen übergeordneten Kategorie nutzt, bleibt ihm überlassen. Das Problem für den Taxonomen besteht nun darin zu entscheiden, mit wievielen Merkmalen eine Gruppe ausreichend definiert ist. Theoretisch würde ein "echtes" apikales Merkmal genügen, dessen Festlegung jedoch aus dem oben gesagten nicht möglich ist. In der Praxis deutet der Taxonom immer mehrere charakteristische Merkmale heraus. Das Ziel sollte dabei zum einen die Herausarbeitung von vielen vorläufig apikalen Merkmalen, zum anderen die stete Überprüfung derselben an neu untersuchten Naturgegenständen innerhalb der gleichen Kategorie sein. Diese Form der Vorgehensweise wird als numerische Taxonomie oder Phänetik bezeichnet (z.B. Sokal & Sneath 1963). Basale Merkmale sind aus der Definition einer Kategorie herauszuhalten, weil sie nicht spezifisch für sie sind. Die Definitionsliste einer Kategorie besteht hiernach aus einer Mischung an quantitativ und qualitativ überprüften und zu überprüfenden Merkmalen, welche allerdings nichts über Komplexität, "Primitivität", "Fortschrittlichkeit" und Verwandtschaft aussagen. |
Hierarchisch geordnet sind im System nur die Merkmale und die dadurch erzeugten künstlichen Kategorien mit ihren abstrahierten Naturgegenständen. Ein "natürliches" System ist also innerhalb dieser Vorgehensweise nicht zu erreichen, trotz anderslautender Behauptung (z.B. Simpson 1967, Mayr 1975, Panchen 1992). Die Frage, ob die imaginäre "natürliche Ordnung" hierarchisch aufgebaut ist, kann somit (und überhaupt, wie später noch dargelegt) nicht beantwortet werden. Eine qualitative Bewertung des Individuums hinsichtlich dessen Stellung im System ist somit illegitim. Aussagen wie "spezialisierte" oder "ursprüngliche" Formen sind selbst innerhalb des abstrahierten Kategorien-Systems höchst spekulativ und übersteigen dessen Aussagefähigkeit. Ordnung wurde vom Taxonomen oder Systematiker erzeugt, sie ist mittels der genutzten Methode auf "die Natur" nicht übertragbar. Die so erzeugte Ordnung hängt vom Standpunkt des Naturforschers und dessen benutzten Ordnungsprinzip ab. Es liegt hier also bereits ein aus der Physik altbekanntes Phänomen vor, das hier als das biologische Unschärfeprinzip bezeichnet wird. Was heißen soll, daß der Naturforscher allein durch sein Dasein und die Art der Beobachtung Einfluß auf den untersuchten Naturgegenstand ausübt. Die Relativität dieses Prozesses bedingt somit u.a. auch die zu beobachtende Schwemme unterschiedlicher Systeme. Der Einfachheit halber und zur Verständigung der Naturforscher untereinander sollte man sich entweder auf ein einziges Prinzip einigen, welches zukünftig Anwendung finden kann, oder man akzeptiert die "Polysystematik" wie sie heute besteht, wobei hier für jeden einzelnen Bearbeiter die Darlegung seines Ordnungsprinzips Grundvoraussetzung sein sollte. |
Sich dieser oben beschriebenen Einschränkungen bewußt, bietet die Taxonomie jedoch eine pragmatische Möglichkeit, die Vielfalt der Naturgegenstände in den Sammlungsräumen so zu ordnen, daß sie jederzeit auffindbar sind. Weiterhin sind alle neu hinzukommenden Naturgegenstände auf ihre Stellung im System überprüfbar, wobei ab und zu einige auch Anlaß zur Modifikation des bestehenden Systems geben. Der praktisch arbeitende Taxonom erarbeitet sich somit prinzipiell einen Bestimmungsschlüssel zur Identifikation der zu bearbeitenden Naturgegenstände. Dies beruht auf der Basis des Ausschlußverfahrens: " Merkmal A bei Naturgegenstand X vorhanden - Bestätigung durch Merkmale B, C, ... - Identifikation von Naturgegenstand X " Merkmal A bei Naturgegenstand X nicht vorhanden - Wechsel zu Merkmal B und Überprüfung an Naturgegenstand X. Die in der Praxis z.T. überaus komplexen Bestimmungsschlüssel gründen sich einzig auf die methodischen Grenzen des Systems (z.B. die Nichtberücksichtigung aller Varianten). Diese Vorgehensweise hat sich für die Bestimmung der heute lebenden Naturgegenstände aus gut untersuchten Kategorien und regionalen Gebieten bewährt. So vertraut doch jeder Biologiestudent heute auf die ausgefeilten Bestimmungsschlüssel für die Naturgegenstände, die er zu bestimmen hat. Für den Taxonomen, der sich an Naturgegenstände einer Terra incognita heranwagt, brechen dagegen schwere Zeiten an. Er muß erst den ganzen Weg wie oben beschrieben durchexerzieren, bevor er dieses Hilfsmittel des Bestimmungsschlüssels zur Verfügung stellen kann. Es fängt mit dem einfachsten Schlüssel an: Naturgegenstand X zeigt Merkmal A, d.h. alle Naturgegenstände mit Merkmal A können vorläufig bestimmt werden als Naturgegenstand X. Zukünftige Vergleiche an anderen Naturgegenständen werden dies verifizieren oder falsifizieren. Der Bestimmungsschlüssel wird stetig erweitert, spezifiziert und korrigiert. Es ist deutlich zu sehen, wie künstlich und willkürlich man vorgeht, ja vorgehen muß. Aussagen über "Natürlichkeit eines Merkmals" oder eines bestimmten Naturgegenstandes werden ad absurdum geführt. Im Bereich der fossilen Naturgegenstände tritt häufig der Fall auf, daß entweder gänzlich vorher unbekannte Naturgegenstände gefunden werden (analog zu Naturgegenständen aus heute noch unerforschten Regionen), oder daß Naturgegenstände zu unvollkommen bzw. nur eingeschränkt untersuchbar vorliegen, z.B. bei bruchstückhafter Erhaltung von Fossilien. Auch hier geht man zwecks Bestimmung von bereits bestehenden Taxa (Kategorien) aus, wobei im Gegensatz zur Rezentsystematik häufig das für Vergleichszwecke benötigte Material aus nur wenigen entsprechenden Naturgegenständen besteht (dies findet man besonders häufig in der Wirbeltierpaläontologie). Die Unsicherheit und Instabilität des im Fossilbereich erzeugten Systems ist damit noch größer als im Rezentbereich. So sind auch die gleichlautenden Kategorien beider Bereiche unter Umständen verschieden definiert (z.B. "Art"; s. hierzu Junker & Scherer 1998). Damit ergibt sich eine enorme Problematik, wenn, wie geschehen, die Systeme der fossilen und rezenten Naturgegenstände zusammengelegt werden bzw., wenn basierend auf dem historisch früher erschienenen rezenten System (z.B. dem von Linné) die fossilen Naturgegenstände dort eingeordnet wurden. Folgen davon sind: Diagnosen von Kategorien, die sich in der Rezentsystematik bewährt haben, werden schwammig; einzelne fossile Taxa brechen bestehende Kategorien auf. Der Taxonom ist also darauf angewiesen, alles verfügbare Datenmaterial, sowohl von fossilen wie rezenten Naturgegenständen, zu verwerten und das bestehende System stetig zu überprüfen und anzugleichen. Voraussetzung ist allerdings, daß er wirklich alle Naturgegenstände einordnen will. Findet diese permanente Überprüfung und Überarbeitung mit all ihren Konsequenzen (z.B. Aufspaltung bzw. Fusion von Kategorien gleichen oder verschiedenen Ranges) nicht statt, dann arbeitet der Taxonom / Systematiker nicht stringent. Es wäre dann sinnvoller, die rezente Systematik von der fossilen zu trennen. Eine weitere evidente Schwierigkeit zeigt sich in der Interpretation von bestimmten Merkmalen als Mischmerkmale. Diese werden nun nicht mehr als unabhängige Charakteristika betrachtet, sondern bewußt oder unbewußt in die Nähe der "Endmerkmale" plaziert, was sich auch in der Stellung innerhalb des Systems zeigt. Die so erzeugten Mischgruppen liefern den Phylogenetikern häufig das fossile Material zum Belege ihrer evolutiven Hypothesen (z.B. Eldredge & Cracraft 1980; s. Kap. "Taxonomie und Phylogenie"). Ein Beispiel wäre das Vorkommen von Federn unter Wirbeltieren. Dadurch, daß Federn als ein "exklusives Merkmal" der Vögel betrachtet worden ist, ist der Nachweis von federähnlichen Strukturen bei kleinen Raubdinosaurier-Skeletten aus China für Phylogenetiker der langersehnte Beleg für die Theorie der Abstammung der Vögel von Raubdinosauriern. Das Merkmal "Vorkommen von Federn" erfährt somit bereits vorab seine Interpretation als "Übergangsmerkmal", was methodisch unzulässig ist. |
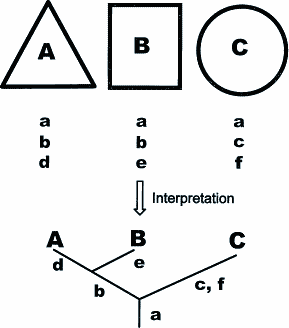
Zwangsläufig werden durch das oben beschriebene Prozedere abgrenzbare Gruppen im System näher oder weiter voneinander positioniert. Grundlage ist das Vorhandensein oder Fehlen von basalen Merkmalen innerhalb der nächst größeren (oder hierarchisch: höheren) Kategorie: Angenommen Kategorie A und Kategorie B stehen im System näher zusammen als Kategorie A und Kategorie C, so bedeutet dies, daß Kategorie A und B mehr basale Merkmale teilen, als dies bei Gruppe A und C der Fall ist (Abb. 1). Der Taxonom liefert die diagnostizierbaren Kategorien, während der Systematiker sie nach der Quantität und Qualität ihrer Merkmale ordnet. Doch keiner von beiden hat basierend auf der Methode seines Erkenntnisgewinnes in irgendeiner Weise die Möglichkeit, naturwissenschaftlich fundierte Aussagen über einen Verwandtschaftsstatus dieser Kategorien untereinander zu treffen. Beispiel: Ein Taxonom hat die Kategorien "Dreiecke", "Quadrate" und "Kreise" durch geometrische Merkmale wie Form, Winkel, Geradlinigkeit etc. definiert. Der Systematiker erstellt daraufhin ein mögliches System in dem er die "Dreiecke" näher bei den "Quadraten" plaziert, die beide gewissermaßen eine eigene Kategorie neben der Kategorie "Kreise" bilden. Begründet werden kann dies durch Merkmale wie Winkelbildung, größere Formähnlichkeit usw. (Abb. 1). Welche der erzeugten Kategorien sind nun näher miteinander verwandt? Vielfach würde diese Frage in diesem Zusammenhang gar nicht gestellt werden, da die Künstlichkeit evident ist. Doch sind - wie oben beschrieben - auch alle naturgegenständlichen Kategorien nichts anderes als künstliche Gebilde, z.B. "Aves (Vögel)", "Dinosauria". Zu fragen wäre also in diesem Zusammenhang: Was bedeutet in der Biologie eigentlich der Begriff "Verwandtschaft"? Es ist gewöhnlich die Feststellung der Position in einem dokumentierten Stammbaum von sich fortpflanzenden Organismen gemeint. Beobachtet und dokumentiert man solch eine Organismen-Gemeinschaft über einen bestimmten Zeitraum hinweg (z.B. Ahnentafeln), so lassen sich über die einzelnen Individuen Aussagen zur verwandtschaftlichen (genetischen) Position treffen. Verwandtschaftliche Aussagen sind explizit aber nur in diesem besonderen, eng umgrenzten Fall möglich. Biologische (genetische) Verwandtschaftsaussagen sind im Fall der Kategorienbildung (Art, Gattung,...) methodisch bedingt nicht möglich, da hier künstlich erzeugte Gruppen vorliegen. Eine Aussage, die jedoch getroffen werden kann, ist: Bei Betrachtung von entsprechenden Merkmalsparametern ist Kategorie A ähnlicher mit Gruppe B, als mit Gruppe C. Dies impliziert aber nicht eine biologische oder natürliche Verwandtschaft. Folgerichtig sollte man also den Begriff der "Verwandtschaft" für alle künstlichen Kategorien fallen lassen. Was man feststellen kann, sind Ähnlichkeiten oder Unähnlichkeiten, was man bildet, sind Ähnlichkeitsgruppen oder Äquivalenzklassen (M. Gutmann 1993). Eine oft unbewußte Vermischung der methodischen Ebenen findet man in der Aussage "Kategorie A ist ähnlicher mit Kategorie B als mit Kategorie C, also ist die Wahrscheinlichkeit einer näheren Verwandtschaft zwischen Kategorie A und B größer als zwischen Kategorie A und C", was somit einen methodisch unzulässigen Rückschluß darstellt. Ein Beispiel wäre das innerartliche Phänomen der Doppelgänger bei uns Menschen. Hier ist zwar phänotypisch eine große Ähnlichkeit festzustellen, die sich aber nicht zwangsläufig auf eine natürlich-genetische Verwandtschaft zurückführen läßt. |
Taxonomie läßt sich also nach all dem bisher Gesagten als Möglichkeit definieren, die natürliche Vielfalt künstlich zu ordnen und systematisch zu erfassen, wobei jedoch keine Aussagen über eine biologische Verwandtschaft möglich sind. Wie sieht es nun mit der Veränderlichkeit der Naturgegenstände im Laufe der Zeit aus? Ist die Taxonomie auch ein Hilfsmittel zur Sichtbarmachung eines Prozesses, der die Naturgegenstände verändert und diese damit immer wieder auf die Bewährungsprobe stellt (Evolution)? |
|
Vorab einige Bemerkungen zu diesem Prozeß der Phylogenese. Diese beschreibt einen historischen Vorgang (Junker & Scherer 1998, Kap. I.1), ist also streng genommen naturwissenschaftlich nicht überprüfbar. Ihr Ablauf ist einmalig, sie ist nicht reproduzierbar. Sie wird somit immer den Rang einer mehr oder weniger gut fundierten Hypothese haben und sie ist nicht mathematisch beweisbar, allenfalls mittels Indizien belegbar (Abb. 3). Diese Indizien lassen verschiedene Deutungen zu, denn die sich damit befassenden Naturforscher (= Natur-Historiker) sind Menschen, die gesellschaftlich und ideologisch eingebunden und somit den kulturellen, gesellschaftspolitischen und religiösen Strömungen ihrer Zeit ausgesetzt sind. Es existieren daher heutzutage drei verschiedene Modelle zur Erklärung der Phylogenese: das evolutionistische, das kreationistische und das theistisch-evolutionistische Konzept. Die Lager ihrer Repräsentanten trennen eminente Grundsätze der jeweiligen Weltanschauung und sind durch Naturwissenschaft alleine nicht überbrückbar. Es bleibt aber die Möglichkeit, Daten und grundverschiedene Ansätze zu verknüpfen bzw. sie in ein bestehendes Weltbild einzupassen (Abb. 4). Dabei gilt: alle drei Modelle sind letztlich nicht naturwissenschaftlich beweisbar. Der Naturforscher entscheidet sich für das eine oder andere aufgrund seiner Lebensphilosophie; sogenannte naturwissenschaftliche Gründe untermauern nur das eigene Weltbild. Als Wissenschaftler sollte man allerdings offen für die Argumentation des anderen Lagers sein, was bedeutet, daß nach Kenntnisnahme der Regeln der Alternativhypothese eine sachliche Beurteilung und eine Nachvollziehbarkeit erwartet werden kann. Dies wird leider heute von allen drei Parteien nur selten in einer akzeptablen Form vollzogen, statt dessen finden Grabenkämpfe statt, die häufig in persönlichen Diffamierungen enden. |
|
Was hat nun Taxonomie hiermit zu tun? Der Taxonom liefert die Daten in Form von abstrahierten Kategorien. Er sollte es allerdings strikt vermeiden, sich bei der Errichtung der Gruppen von phylogenetischen Standpunkten leiten zu lassen. Sonst werden hierbei zwei Methodenebenen miteinander vermischt, die Aussagefähigkeit würde zwangskanalisiert. Es kommt zum wissenschaftstheoretisch zu vermeidenden Zirkelschluß. Folgendes, bereits weiter oben angeführtes Beispiel soll dies erläutern. Ein Paläontologie findet ein Dinosaurierskelett mit Federn. Er wertet das Merkmal "Federn" so stark, daß hier nach der vorgefaßten Theorie der Abstammung der Vögel von Dinosauriern, eine Mischform vorliegt, die die Theorie wiederum belegt. Ein oft publiziertes Beispiel eines wissenschaftstheoretischen Zirkelschlusses. Was der Phylogenetiker mit den vom Taxonomen erzeugten Kategorien macht, ob er sie evolutionistisch oder kreationistisch interpretiert, braucht den Taxonomen nicht zu berühren, sie müssen bei ihrer Errichtung nur frei vom phylogenetischen Gedankengut sein. |
Diese von Hennig (1950) begründete Methode hat den Anspruch, verwandtschaftliche Verhältnisse von Naturgegenständen durch Bewertung von Merkmalen als apomorph (abgeleitet) bzw. plesiomorph (ursprünglich) aufzuzeigen (z.B. Wiley 1981, Smith 1994). Dabei gilt bei der Verzweigung des kladistischen Stammbaumes das Prinzip der Dichotomie (= Gabelung; Abb. 5). Zum Kladismus sind zwei große Kritikpunkte anzumerken. Zum einen ist immer mehr festzustellen, daß hierbei eine "Einmischung" der phylogenetischen Interpretation in die taxonomische Ebene stattfindet. Diagnosen von Kategorien werden "ergänzt" mit Merkmalen, die nach der phylogenetisch-systematischen Methode den Anspruch auf verwandtschaftliche Aussagekraft besitzen. So werden notwendigerweise Merkmale von heute lebenden Organismen in die Diagnosen aufgenommen, die in der Praxis des Fossilienforschers kaum Nutzen haben, da sie nur selten am fossilen Material überprüfbar oder in ihrer Ausprägung und Konstanz zu variabel sind (Variabilität und Konvergenz). So bietet sich auch hier wieder das Merkmal "Vorkommen von Federn" als Beispiel an. Vom kladistischen Standpunkt her ist es eben nicht nur ein X-beliebiges Merkmal, sondern verbindet von vornherein zwei Kategorien hierarchisch miteinander: Die "einfachen" Federn der Raubdinosaurier wären so die Vorstrukturen der "komplexen" Vogelfedern. |
|
Der zweite Kritikpunkt betrifft die vermeintliche Aussagekraft des erzeugten Kladogramms in Hinblick auf verwandtschaftliche Fragen bezüglich der geordneten Kategorien. Wie bereits von M. Gutmann (1993) dargelegt, sind Kladogramme nichts anderes als eine weitere Klassifizierungsform nach Maßgabe von diskreten Merkmalsparametern. Die so geordneten Systeme haben dieselbe Aussagekraft über verwandtschaftliche Verhältnisse der einzelnen künstlichen Kategorien untereinander, wie die der klassischen Schemata - keine! Was hier geordnet wird sind - analog zu dem oben beschriebenen - Ähnlichkeitsgruppen. Somit liefert der kladistische Ansatz eine weitere Klassifizierungs- und Darstellungsform. Sich der Grenzen dieser Methode bewußt bietet sie aber eine mehr oder weniger übersichtliche Möglichkeit, die Naturgegenstände zu ordnen. Allerdings sollte dabei die Vermischung mit den Systemen der klassischen Taxonomie vermieden werden, da ansonsten ein Wirrwarr an Kategorie-Bezeichnungen beider Methoden im Umlauf wären. Im ungünstigsten Fall hätte man gleichlautende Kategorie-Bezeichnungen, die aber unterschiedlich definiert wären. |
Der Taxonom greift durch seine ordnende Tätigkeit direkt (Entnahme von Naturgegenständen) und indirekt ("Merkmalsfindung" = Merkmalsproduktion zu Diagnosezwecken) in die Natur ein. Dies hätte zweifelsohne Auswirkungen auf eine natürliche Ordnung, vorausgesetzt sie existiert. Da der Taxonom ein Teil der Natur ist, unterliegt er auch ihren "Ordnungen" (Naturgesetzen). Es ist ihm dadurch unmöglich, von außen, d.h. objektiv an diese Problemstellung heranzugehen, ganz abgesehen von seiner relativen Position in der Betrachtung des Naturgegenstandes selbst, wobei die Schwerpunkte seiner Betrachtung kulturell und intuitiv beeinflußt werden. Dies bedingt die beiden folgenden Sichtweisen, wie man an die Beschreibung von Naturgegenständen herangehen kann: 1. Man beschreibt einen Naturgegenstand entweder in seiner natürlichen oder naturgemäßen Umgebung (Kulturlandschaft; "natürliche" Laborbedingungen), ohne diese Daten für taxonomisch-systematische Zwecke zu nutzen. 2. Man betrachtet den Naturgegenstand nur zu Ordnungszwecken und abstrahiert ihn entsprechend. Es liegt hier ein dualistisches Prinzip vor. Beide Betrachtungsweisen sind legitim, schließen sich aber in ihrem Ergebnis gegenseitig aus. Ein Naturgegenstand "natürlich ordnen" zu wollen, ist somit ein Widerspruch in sich. Abstufungen zwischen diesen beiden Positionen sind möglich - es kommt zum Phänomen des biologischen Unschärfeprinzips (s. Kap. Systematik und Hierarchie). Je mehr der Naturforscher sich mit den Naturgegenständen beschäftigt, desto weniger "natürlich" sind die Ergebnisse. Die künstlich erzeugte Ordnung verhindert eine natürliche Beobachtung oder Bearbeitung. Doch selbst wenn man Naturgegenstände ausschließlich in ihrer natürlichen Umgebung ohne ordnende Hintergedanken beobachten und beschreiben würden, hätten wir niemals die Möglichkeit, die imaginäre Naturordnung erkennen zu können, da wir ja Teil dieser "Naturordnung" selbst sind und somit den notwendigen Standpunkt des Außenstehenden niemals einnehmen können. Da es jedoch allein aus pragmatischen Gründen sinnvoll ist, die Naturvielfalt zu ordnen, ist die Legitimation von künstlichen Systemen gegeben. Man sollte sich nur davor hüten, sie als Abbildungen einer "Naturordnung" oder als Abbildungen einer Phylogenese zu interpretieren. Als Teil von dem Ganzen "Natur" steht der Naturforscher auch in Kontakt mit den anderen Teilen, also eben jenen Naturgegenständen, die er beschreiben will. Dadurch, daß er auf die oben beschriebene Weise in "die Natur" eingreift, muß das damit auch Auswirkungen auf ihn selbst und vielleicht auch auf die von ihm benutzte Methode haben. Sind diese Auswirkungen real beobachtbar und wie können sie aussehen? Ist die "Naturordnung" durch die Tätigkeit der Naturforscher verändert worden? Daß der Mensch Auswirkungen auf die Naturlandschaft ausübte und ausübt steht außer Frage: In Mitteleuropa gibt es überwiegend nur noch Kulturlandschaften. Wenn man allerdings von einer ursprünglichen Naturlandschaft spricht, meint man eine Umgebung, die ohne Einfluß der Kategorie Homo sapiens geblieben ist. Dies impliziert aber, daß die Veränderungen an einer Landschaftsstruktur, die von anderen Naturgegenständen ausgehen (z.B. bei der Erstbesiedlung einer vulkanisch entstandenen Insel durch Pflanzen und Insekten oder dem Weideverhalten von großen pflanzenfressenden Säugetieren), hier "natürlich" genannt werden muß. Dies belegt man mit der Tatsache, daß nur der Kategorie Homo sapiens Kultur zugebilligt wird. Die anderen Naturgegenstände hätten somit nicht die Möglichkeit, sich dessen bewußt zu sein, was sie sind und tun und das, was sie tun, im voraus zu planen. Doch wer von uns kann entscheiden, ob dies wirklich so ist? Dank: Ohne die Diskussionen mit den folgenden Kollegen, wäre dieser Artikel nicht möglich gewesen: Dr. D. Cook (Zürich), Dr. M. Gutmann (Frankfurt), Dr. B. Herkner (Karlsruhe), M. Verhaagh (Karlsruhe). Auch für deren kritische Durchsicht des Manuskriptes sei ihnen und Dr. R. Junker (Baiersbronn), Frau Dr. J. Fehrer und Dr. H. Ullrich herzlich gedankt. SDG. |
Glossar
|
Literatur
|

|