|
|
|
|
Die Evolutionstheorie geht davon aus, daß durch das Zusammenwirken von zufälliger Mutation und anschließender Selektion nicht nur eine Differenzierung und Spezialisierung bereits vorhandener Strukturen erfolgen kann (Mikroevolution), sondern neue, vorher nicht vorhandene Konstruktionen entstehen können (Makroevolution). Es wird davon ausgegangen, daß evolutionäre Veränderungen lange Zeiträume benötigen, was zum Teil an der relativen Seltenheit von Mutationen liegt. Umgekehrt kann auf dieser Basis angenommen werden, daß eine Erhöhung der Mutationsrate eventuell zusammen mit einer stärkeren Selektion Evolution beschleunigen und damit eventuell im Laborsystem beobachtbar machen könnte.
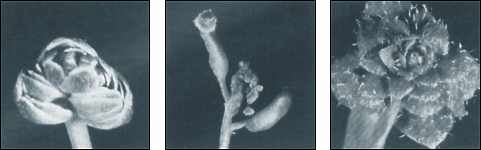
Künstliche Mutagenese ist ein wichtiges molekularbiologisches Verfahren, um Aussagen über die Funktion von Proteinen oder Genen zu gewinnen. Dabei wird künstlich die Mutationsfrequenz beispielsweise in Pflanzensamen erhöht und die so gewonnenen Mutanten nach interessierenden Mutationen durchsucht, welche dann weiter untersucht werden. In der Mutagenese finden Röntgenstrahlen, schnelle Neutronen und Chemikalien wie Ethylmethansulfonat (EMS) Verwendung. EMS führt vornehmlich zu spezifischen DNA-Veränderungen (Alkylierung), wodurch bei der Replikation das Basenpaar von G:C zu A:T verändert wird (Sega 1984, Burns et al. 1986). Das besondere daran ist, daß EMS vor allem Punktmutationen erzeugt. Extrapolationen von bei Mikroorganismen gefundenen Raten ergeben für Arabidopsis eine abgeschätzte Spontanmutationsrate von 0.0033 pro Genom und Replikation (Drake 1991, Drake et al. 1998), wobei einzelne Genorte des Arabidopsis-Genoms recht unterschiedliche Raten aufweisen können. Eine Studie fand für genetisch bestimmte Merkmale, die mit der Selbstreproduktion in Zusammenhang stehen wie die Anzahl der Früchte oder die Anzahl der Samen pro Früchte eine im Verlauf mehrerer Generationen um einen Mittelwert schwankende Variabilität (Shaw et al. 2000). Bei Arabidopsis wird bevorzugt Mutagenese mittels EMS betrieben, da dieser farblose, leicht flüchtige, bei Zimmertemperatur flüssige Stoff ein leistungsfähiges Mutagen ist, welcher zugleich nur eine geringe Sterblichkeit verursacht. Dabei werden jeweils einige Tausend Arabidopsis-Samen in einer 100 millimolaren EMS-Lösung einige Stunden gebadet und anschließend kultiviert. Unter EMS ist die Mutationshäufigkeit um ein vielfaches erhöht (Koorneef et al. 1982) - wahrscheinlich ein Faktor der Größenordnung einige 1000. In der 1. Generation mutagenetisch veränderter Pflanzen kommen typischerweise chlorophyllarme Areale in geringer Anzahl (eine Pflanze unter 100 bis 1000) vor.2 Ihr Auftreten gilt als Indikator für eine erfolgreiche Mutagenese, ihr Auftreten erklärt sich wie folgt: Pflanzensamen sind vielzellige Organe. Durch Mutagenese werden in einigen ihrer Zellen genomische Veränderungen erzeugt, in anderen nicht. Es entstehen so adulte Pflanzen, die Mosaike aus intakten und veränderten Genomen sind, was sich beispielsweise in dem Vorliegen einzelner chlorophyllarmer Blätter an einer ansonsten unauffälligen Pflanze zeigt. In der 2. Generation zeigen 20-30% der Pflanzen ontogenetische oder physiologische Veränderungen wie ein allgemein kränkliches Aussehen, vorzeitiges oder verspätetes Blühen, Sterilität oder Zwergwuchs (Reiter et al. 1997) und Chlorosis (Bleichheit durch Chlorophyllmangel). Festzuhalten ist, daß eine EMS-Behandlung durchweg und typischerweise defizitäre Pflanzen hervorbringt. Jede Arbeitsgruppe durchsucht die entstandenen Mutationen nach den für die angestrebten Forschungsfragen relevanten Mutationen. Die weitaus größte Zahl ist jeweils uninteressant und wird verworfen. Aus diesem Grunde existiert keine genaue Aufstellung über alle bisher beobachteten Mutationen. Freilich geben die bisher näher untersuchten Mutationen einen guten Eindruck für typische durch Mutagenese induzierte Veränderungen. So wurde die Bildung anomaler Blüten mit zufälligem Verlust von Blütenorganen beschrieben: Blütenblätter, Kelchblätter, Stempel oder Staubblätter können fehlen (Abb. 2). Ebenso kann die Pflanze eine gestörte Pflanzenorganentwicklung zeigen (Roe et al. 1993, Irish & Sussex 1990, Bowman et al. 1992). Die Zuckerkomposition der Monosaccharidseitenketten von Proteinen der Zellwände kann verändert sein: eine Arbeit beschrieb beispielsweise eine 50 prozentige Reduzierung des Arabinosegehaltes aufgrund eines Defektes in der UDP-L-Arabinose de novo-Synthese (Burget & Reiter 1999). |
|
Es wurden Pflanzen mit einem vorzeitigen Abbruch der Pollenentwicklung, einem Defekt beim Auswachsen der Pollenschläuche oder in der Bildung des Pflanzenembryos beschrieben (Grini et al. 1999). Pflanzen, die homozygot für EMS-induzierte Mutationen sind, können Verwachsungen von Organen zeigen: Beispiele sind Fusionen zwischen Blütenorganen wie Kelchblättern und Blütenblättern ohne Einfluß auf die Fruchtbarkeit oder Fusionen zwischen Eizellen mit nachfolgender Sterilität oder Semisterilität der weiblichen Blütenorgane (Lolle & Hsu 1998). Einige vorteilhafte Mutanten sind ebenfalls beschrieben worden: Arabidopsis-Pflanzen mit vergrößerter Resistenz gegenüber mikrobiellen Pathogenen, beispielsweise verstärkter lokaler Apoptose von Pseudomonas syringae infizierten Zellen mit dem Effekt der Begrenzung der Infektausbreitung (Rate et al. 1999), eine erhöhte Anzahl von Früchten und Samen pro Frucht (Shaw et al. 2000). Diese Neuerungen verbleiben jedoch stets im mikroevolutiven Rahmen. Die durch EMS vornehmlich bewirkte erhöhte Frequenz von Punktmutationen (Transition von G:C zu A:T) ist nur einer von mehreren relevanten Mutationsmechanismen. Weitere Mechanismen sind unter anderem Genumordnungen, Duplikationen, Deletionen oder Inversionen. Die dadurch ausgelösten größeren Veränderungen sind aller Wahrscheinlichkeit nach deshalb aber nicht positiver. Bleibt abzuwarten, ob die symetrische Mutationseffektverteilung, die von Shaw et al. (2000) beobachtet wurde, sich langfristig bestätigt. Die große Anzahl der hier vorgestellten Studien, die Funktionsverluste beschreiben, scheint dies jedoch nicht gerade nahezulegen: Sobald das adaptive Potential ausgeschöpft ist, dürften sich wohl auch hier mehr negative als positive Mutationen zeigen. Hinweise auf Makroevolution sind jedenfalls aus den hier dargestellten Ergebnissen nicht zu gewinnen, aber sehr wohl zu mikroevolutivem Verlust von Funktion und Pluripotenz. |
Anmerkungen
|

|