|
|
|
|
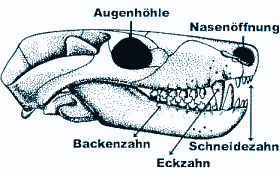
Die 22 heute lebenden Krokodilarten aus den subtropischen und tropischen Regionen unserer Erde gelten gemeinhin als morphologisch uniform. Bei genauerer Untersuchung fallen aber doch diverse Unterschiede auf, die eine Gliederung in drei Gruppen rechtfertigen: 1. die Alligatoren und Kaimane; 2. die echten Krokodile und Sunda-Gaviale; 3. die Gaviale (oder richtiger: Ghariale). Sie unterscheiden sich vom Skelettbau hauptsächlich in der Schnauzenform, der Anzahl und Größe der Zähne, im Bau des Zungenbeinkörpers und in einigen Merkmalen der ersten Halswirbel (Wettstein 1937, Trutnau 1994). Daß im Fossilbericht eine noch wesentlich größere Vielfalt an Arten und Formen repräsentiert ist, ist nur sehr ungenügend bekannt. Die mit diesem Bericht beginnende Serie möchte das ungeheure Spektrum an Formen, Größen und Arten von fossilen Krokodilen vorstellen. Die Bezahnung von heutigen Krokodilen erscheint auf den ersten Blick homodont, d.h. die Zahnkronen der Kiefer sind einheitlich gebaut. So denkt man in puncto Krokodilzähne am ehesten an spitzkonische Formen, ähnlich den menschlichen Eckzähnen. Dies ist eine grobe Vereinfachung und nach heutigen Erkenntnissen in dieser Absolutheit unzulässig. So besitzt z.B. das afrikanische Stumpfkrokodil (Osteolaemus tetraspis) in seinem hinteren Kieferteil halbkugelförmige Knackzähne, und auch das gut bekannte Nilkrokodil (Crocodilus niloticus) hat im hinteren Kieferteil seitlich komprimierte und in seitlicher Ansicht halbkreisförmige Zahnkronen (Trutnau 1994). Doch staunte ein Team US-amerikanischer Paläontologen nicht schlecht, als sie in den Sedimentschichten der Unteren Kreide von Malawi (Ostafrika) typische Backenzähne eines mutmaßlichen Säugetiers mit Krokodilresten zusammen fanden. Clark et al. (1989) konnten daraufhin zeigen, daß hier tatsächlich eine erdgeschichtlich frühe Krokodilart spezialisierte, mehrhöckrige Zähne in der hinteren Kieferregion aufwies (Abb. 1). |
|
Spätere Bearbeiter wiesen diese Mischung an Merkmalen auch bei anderen zeitgleichen Formen aus China nach (Wu et al. 1995). Letztere vergaben aufgrund der Mixtur an Merkmalen aus den Wirbeltiergruppen Crocodilia und Mammalia (Säugetiere) diesen Funden den Namen Chimaerasuchus paradoxus. Auch hier war die herausragende Besonderheit das Vorkommen von mehrhöckrigen "Backenzähnen" im hinteren Teil der Kieferregion. Wu et al. (1995) spekulierten sogar darüber, ob diese Krokodilart nicht gänzlich auf Fleisch als Nahrung verzichtet habe und sich rein pflanzlich ernährte. Diese Folgerung basiert allerdings einzig auf der besonderen Zahnform und ist ohne belegten fossilen Mageninhalt oder ausgeschiedenen Nahrungsresten (Koprolithen) nicht schlüssig nachzuvollziehen. Gomani (1997) beschrieb die Krokodilreste aus Malawi erneut und benannte sie als Malawisuchus mwakayungutiensis im Hinblick auf den Fundort. Die größte Ähnlichkeit mit anderen Krokodilen fand Gomani im Vergleich mit einer nur fossil bekannten Krokodilfamilie - die Notosuchiden, eine fast ausschließlich auf die Südkontinente beschränkte Gruppe. Gomani stellte fest, daß Malawisuchus mit einer Schädellänge von maximal 10 cm und einer geschätzten Maximallänge des Körpers von 60 cm ein sehr kleiner Vertreter einer überaus spezialisierten Krokodil-Gruppe war. Die "Backenzähne" von Malawisuchus sind nach Clark et al. (1989) und Gomani (1997) Anzeichen einer beginnenden Zerkleinerung der Nahrung durch das Gebiß, wie man sie in dieser Art und Weise von den heutigen Krokodilen nicht kennt. Weitere Besonderheiten stellten die Autoren mit dem Überbiß (vgl. Abb. 1) und dem Bau des Kiefergelenks fest, welches neben einer scharnierartigen Auf-und-ab-Bewegung auch eine Bewegungsrichtung nach vorne und hinten zuließ. Daraufhin rekonstruierte Gomani (1997) die Kiefermuskulatur von Malawisuchus derart, daß die Autorin auf ein Beutespektrum aus Insekten, Amphibien und Schnecken folgerte. Weiterhin weist der schlanke Bau der Extremitäten laut Gomani (1997) auf eine rein terrestrische (landlebende) Lebensart hin, also entgegen vieler heutiger amphibisch lebender Krokodile. Eine besondere Annahme lieferte Gomani (1997) mit der Vorstellung, daß Malawisuchus eine grabende Lebensweise besessen haben soll. Hinweise dafür sah sie im Bau der Hals- und Schädelregion, welche die Rekonstruktion einer kräftigen Halsmuskulatur zuließ. Außerdem sollen die sedimentären Strukturen der Fundschicht und die Lage der Fossilexemplare verschüttete Grabgänge belegen. Damit wäre Malawisuchus eine kleinwüchsige, kurzschnauzige Krokodilform, die vorwiegend mit dem Kopf vorwärts grabend in der Nähe von Flussläufen lebte. Die Argumentation scheint auf den ersten Blick schlüssig, doch müssen die Ergebnisse durch zukünftige detailliertere Untersuchungen zur Biomechanik dieser Krokodil-Konstruktion noch abgesichert werden.
Wie ist aber zu erklären, daß in einer Krokodilfamilie wie den Notosuchiden, die gemeinhin als ein Zweig der "primitiven" Vorkrokodile betrachtet wird (Benton & Clark 1988, Clark 1994) so spezialisierte und - nach der Argumentation der Kladisten - fortschrittliche Formen entstanden sind? Die von Clark et al. 1989, Wu et al. 1995 und Gomani 1997 angefertigten kladistischen Analysen zum Aufzeigen von "Verwandtschaften" zwischen Malawisuchus und ähnlichen Krokodilen verwirren in dieser Hinsicht angesichts der Vielzahl an vorgestellten "Stammbäumen" (Kladogrammen) nur (Abb. 3). Leider wird methodisch auch in diesen wichtigen Arbeiten der Schwerpunkt auf die phylogenetisch-systematische Interpretation gelegt, wobei stets das Ziel war, Verwandtschaftsbeziehungen zu ermitteln. Nach Herkner (1999) und Rossmann (in Vorbereitung) ist diese Interpretation methodisch stark zu hinterfragen. Die Frage nach den Zusammenhängen zwischen den verschiedenen Krokodiltypen der Kreidezeit bleibt letztlich ungeklärt, wie auch der Grund nach dem offensichtlichen Verschwinden dieser interessanten Gruppe zum Ende der Unteren Kreide hin. |
|
|
Literatur
|

|