| Merkmalskonflikte und phylogenetische Erwägungen
|
Ostafrika bekommt Ahnen-Konkurrenz:
"Little Foot" und sein Skelett
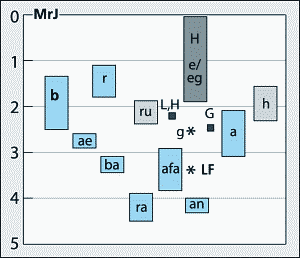
Ostafrika mit seinen reichen Funden hat in Sachen "letzter direkter Vorfahre" des Menschen meist die alleinige Vorherrschaft. Dazu gehören der 2,9-3,8 MrJ alte A. afarensis (das Skelett war als "Lucy" bekannt geworden), der 4,4 MrJ alte A. ramidus, der 4,2 MrJ alte A. anamensis oder der neue Fund A. garhi. Doch inzwischen meldet sich sein Konkurrent Südafrika, der erste Fundort früher Hominiden, wieder mit neuen und aufregenden Funden zu Wort. Bis jetzt kannte man von dort A. africanus (2,5-3 MrJ) als relevanten Vorfahren.
Für das 1995 als "Little Foot" veröffentlichte Fußskelett (Clarke & Tobias 1995), das zu einem vor über 3 MrJ in Südafrika lebenden Hominiden gehörte, fand sich nämlich letztes Jahr ein ansehnlicher Teil seines Restskelettes. "Little Foot" ist teilweise ein sogenannter Museumsfund, der bei Revisionsarbeiten in verschiedenen Museumsschubladen der Universität von Witwatersrand, Südafrika, wiederentdeckt und als Hominide klassifiziert wurde. Weitere Knochen wurden in anderen Schubladen gefunden, bis Ron Clarke zielsicher an der Fundstelle neue Grabungen durchführte, um den "Rest" zu bergen. Das Team war erfolgreich; neben einigen Langknochenfragmenten fand man Schädel und Unterkiefer, was alles zu dem gleichen Skelett gehörte, inzwischen bekannt unter der Katalognummer StW 573 (Sterkfontein West, 3,2-3,6 MrJ). Das Skelett gehört wohl zu einer Australopithecus-Art, unter Umständen zu A. afarensis (wobei dieser dann eine wesentlich südlichere Verbreitung gehabt hätte als bisher angenommen) oder A. africanus. Sowohl A. africanus als auch der neue Fund A. spec. besaßen nach neuesten Ergebnissen ein affenähnlicheres Skelett als A. afarensis (siehe auch Hartwig-Scherer 1998, Oliwenstein 1995) und die Debatte um das Wie und Wann der Evolution zum obligat aufrecht gehenden Menschen bleibt spannend. Damit erhielt Südafrika erneuten Aufschwung im Wettrennen um den letzten direkten Vorfahren des Menschen.
|
|
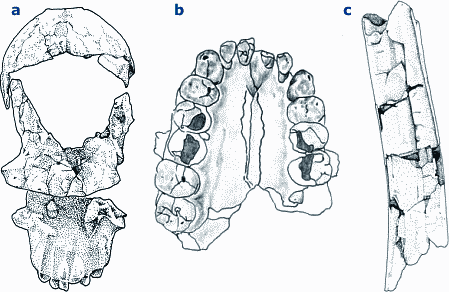
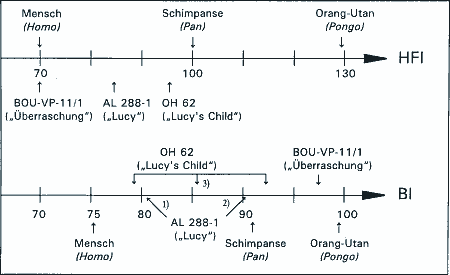
Die neue Art füllt eine klaffende Lücke zwischen 3 bis 2 MrJ, eine Zeit, in der Ostafrika, die hypothetische Wiege der Menschheit, keinen Kandidaten für den Übergang zwischen dem immer-noch-Favoriten A. afarensis und dem inzwischen reklassifizierten "Homo" = Australopithecus habilis aufweist. Ein solches Szenario jedenfalls scheint Tim White zu implizieren, wenn er A. garhi als möglichen direkten Vorfahren des Menschen in die Fachwelt einführt. Er spricht von A. garhi als einer Übergangsform zwischen einem "zweibeinigen Schimpansen" mit großen Zähnen und Aasverzehr zu einem großhirnigen fleischvertilgenden Menschen. Dieser Übergang soll in weniger als 500.000 Jahren stattgefunden haben, was für einige Anthropologen viel zu wenig Zeit ist (Culotta 1999). Neben dem menschlich anmutenden Beinskelett hat er jedoch den "Nachteil" eines überaus ursprünglichen Schädels und sehr affenähnlich anmutender Vorderextremitäten (Abb. 1). Ein ernsthaft diskutierter Kandidat für diese Position bleibt weiterhin der mehr oder weniger zeitgleich lebende A. africanus, dessen wesentlich grazilerer und weniger "primitiver" Schädelaufbau etwas besser für eine solche Position paßt. Allerdings besitzt dieser ein durchgängig recht affenähnliches Skelett, das ihn zwar einerseits mit dem späteren OH62-Skelett ("Homo" = Australopithecus habilis) verbindet, nicht jedoch mit der vergleichsweise "fortschrittlicheren", aber älteren "Lucy". Der Bewertungskonflikt ist offenkundig. Schon früher hat man sich die Frage gestellt, ob man bei der Einordnung eines Fundes nach dem Schädel oder nach den Langknochen gehen soll (Hartwig-Scherer 1998). Das Merkmalsmosaik, das diesen Konflikt verursacht, schafft also "Spielräume" in der phylogenetischen Bewertung, sodaß die jeweilige Wahl oft so auszufallen scheint, daß der gerade diskutierte Fund der "letzte direkte Vorfahre" wird.
Betrachtet man die gesamte Gruppe der Australomorphen, läßt sich kein kontinuierlicher Trend vom Klettern in Bäumen zur Bodenanpassung mit aufrechtem Gang feststellen. Ausgeprägtes Klettervermögen findet sich sowohl bei frühen als auch bei späten Formen. Es wurde sogar bei einer Form identifiziert, die bis vor kurzem noch der Gattung Homo zugeordnet wurde ("Homo" habilis, nach Wood [1999] jetzt Australopithecus habilis; siehe auch den Beitrag über "Homo" habilis in dieser Ausgabe). In dieser Eigenschaft glich dieses Kletterskelett OH62 einem Teilskelett der 3-2 MrJ alten Art A. africanus (McHenry 1996), die ebenfalls als Bindegliedsanwärter diskutiert wird. Die - sehr ambivalent geführte - Diskussion zu A. afarensis (3,8-3 MrJ) legt für "Lucy" ebenfalls ein bestimmtes Maß an Klettern in Kombination mit einer speziellen Form des zweibeinig aufrechten Gehens nahe, während die 4.4 MrJ alte A. anamensis-Form als zweibeinig beschrieben wurde (siehe jedoch Einwände in Junker & Scherer 1998, 257), sodaß kein chronologischer Trend festzustellen ist: Das Motto "ältere Formen klettern mehr als jüngere" kann nicht ohne Ausgliederung vieler anderer Formen aufrechterhalten werden - was wiederum eine phylogenetisch sinnvolle Anordnung der Schädel (Gehirn- und Gesichtsevolution) verhindern würde. Angesichts dieser Tatsache schwindet einiges von der Bedeutung als "Übergangsform", die die Beschreiber dem Teil-Skelett von A. garhi beimessen.
Mehrere frühere Originalpublikationen (Hartwig-Scherer 1991, Korey 1990), die die affenähnlichen Proportionen der späteren habilis-Form aus Olduvai (OH 62) demonstrierten, werden von den Autoren nicht zur Kenntnis genommen, beziehungsweise ungerechtfertigterweise als falsch disqualifiziert: Asfaw und Kollegen bezichtigen McHenry (1998) und Hartwig-Scherer (1991) der Veröffentlichung eines irreführenden und unkorrekten Index, mit dem die Arm-Bein-Proportion des habilis-Skelettes als affenähnlich quantifiziert wurde. Dabei jedoch haben sie übersehen, daß wir diesen Index aus einer Arbeit zitieren, die ebendiese Autoren Asfaw, White, Suwa in einem früheren Artikel selbst mitverfasst hatten (Johanson 1987).
Es irritiert, wenn gegenläufige Indizien nicht nur nicht zur Kenntnis genommen, sondern sogar ungerechtfertigterweise disqualiziert werden, nur weil diese nicht in das erfolgversprechende Phylogenese-Schema passen. Es scheint verlockender zu sein, Hypothesen über Zwischenglieder und potentielle letzte direkte Vorfahren des Menschen zu publizieren, als eine weitere Art der großen Radiationsgruppe der Australomorphen zu beschreiben. Nach Fred Grine (Culotta 1999) besitzt A. garhi nur äußerst wenige Merkmale, die ihn mit Homo verbinden. Damit sei er sicher kein besserer Kandidat für eine direkte Vorfahrenschaft als der südafrikanische A. africanus (siehe auch Kasten). Nach Bernard Wood (Culotta 1999) könnte er genauso gut eine evolutive Sackgasse darstellen. Aus der Perspektive der Grundtypenbiologie ist er ein weiterer Hinweis für die extensive Radiation der Australomorphen, die möglicherweise einen Grundtyp bilden. Wenn diese Form tatsächlich Orang Utan-ähnliches Schwinghangeln mit einem aufrechten Gang verband - falls die beobachteten Proportionen so gedeutet werden können - wäre diese Weite an Merkmalskombinationen tatsächlich eine Überraschung.
| |