|
|
|
|
|
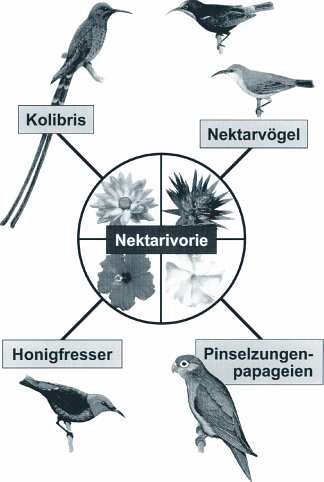
Als Nektarivorie bezeichnet man die Ernährung von Blütennektar, wie sie bei allen Kolibritaxa anzutreffen ist. Diese Ernährungsform ist nur bei wenigen anderen tropischen Vogelfamilien (z.B. Afrika/Asien: Nektarvögel, Nectariniidae; Asien/Australien: Honigfresser, Meliphagidae; Pinselzungenpapageien, Psittaglossidae; Hawaii: Kleidervögel, Drepanididae; Abb. 1) zu finden. Bezüglich der Nahrungswahl sind Kolibris als eutrop zu bezeichnen, d.h. von allen nektarivoren Vögeln stellen sie die spezialisiertesten Blütenbesucher dar, obwohl sie sich außerdem von Insekten, Pollen, Früchten und Baumsäften ernähren. Aufgrund ihres außerordentlichen Artenreichtums und ihrer weiten Verbreitung, verbunden mit zahlreichen gattungs- und artspezifischen Spezialisierungen, kommt den Trochiliden neben den Insekten in der Neuen Welt eine wichtige Bestäuberfunktion zu. Außer den vielfältigen geeigneten anatomisch-morphologischen Konstruktionen des Vogels (vgl. Weller 1999) spielen Blütenerkennung und Blütenmorphologie als wechselseitige Voraussetzungen eine wesentliche Rolle. Hieraus und aus der Konkurrenzsituation der Blütenbestäuber ergeben sich zudem Hinweise auf Fragestellungen nach der Herkunft und Evolution der Nektarivorie, vor allem auch innerhalb der Familie Trochilidae.
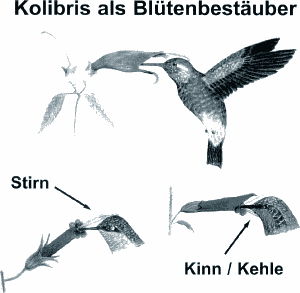
Im Gegensatz zu Insekten, die Blütenfarben lediglich im UV-Bereich erkennen, werden Vögel von Spektralfarben mit leuchtenden Farbtönen angelockt. Von Kolibris besuchte Blüten zeichnen sich vorwiegend durch kräftige Rottöne oder Orange, seltener durch Grün, Blau, Weiß oder Farbkombinationen (sog. Papageienfarben) aus. Untersuchungen an südamerikanischen Pflanzenfamilien, z.B. Gesneriaceen, deuten an, daß außer Blüten auch entsprechend gefärbte vegetative Merkmale wie rote Blattspitzen oder -stengel erkannt werden, um die Auffälligkeit ornithophiler Pflanzen unter den relativ lichtarmen Bedingungen des tropischen Regenwaldes zu erhöhen (Wittmann, mündl.). Der bei Vögeln weitgehend reduzierte bis fehlende Geruchssinn widerspiegelt sich in der Geruchlosigkeit vieler Vogelblumenblüten, was "illegale" Nektarivorie, beispielsweise von Insekten oder Fledermäusen, die keine Bestäubung leisten, ausschließen soll. Wie bereits Johow (1900) bemerkte, ist es in evolutionsgeschichtlicher Hinsicht allerdings nicht erwiesen, ob es sich dabei um eine koevolutive Anpassung handelt, d.h. ob z.B. insektenblütige Pflanzen als potentielle Vorgänger mit der Entwicklung der Ornithophilie auch ihren Geruch verloren haben. Weitere morphologische und physiologische Merkmale der Vogelblumen unterstreichen die enge Beziehung zu ihren Bestäubern (vgl. Faegri & van der Pijl 1966). Gegenüber Insektenblumen ist die Nektarproduktion erheblich gesteigert, was bereits als Indiz auf den Bestäubertyp gelten kann. Die Nektarkonzentration gewährleistet keine ausreichende Versorgung des Kolibris, der dadurch gezwungen ist, weitere Blüten - bis zu 3000 pro Tag - aufzusuchen und damit Fremdbestäubung zu gewährleisten. Die je nach Pflanzentyp und Region mehr oder weniger ausgeprägte Periodizität der Nektarabgabe widerspiegelt sich auch in den Strategien des Nahrungssuchverhaltens der Kolibris. Während einige Arten streng territorial sind und Nahrungsreviere, d.h. beispielsweise blühende Bäume, gegenüber Artgenossen und anderen Kolibris, aber auch Großschmetterlingen und Bienen verteidigen, zeigen andere ein nichtterritoriales Umherstreifen, wobei mehr oder weniger gezielt und regelmäßig bestimmte Futterpflanzen angeflogen werden ("trap lining"). Zu den territorialen Arten zählen die meisten Vertreter der Unterfamilie Trochilinae, während die Phaethornithinae überwiegend als "trap-liner" bezeichnet werden können. Zahlreiche Besonderheiten lassen sich im Bau und der Positionierung der Blüten ornithophiler Pflanzen erkennen. Um ein Abfließen des Honigs zu verhindern, finden sich vielfach Kapillaren, Härchen oder stark benetzbare Oberflächen an Blütenboden und -saum. Die Blüten sind häufig in verlängerten, relativ dichten freistehenden oder -hängenden Blütenständen angelegt. Der relativ derbe Bau der Blüten und ihrer Stengel wird als Anpassung an die erhöhte mechanische Belastung durch die exponierte Position und den Vogelschnabel interpretiert. Im Gegensatz zu anderen nektarbesuchenden Vögeln können Kolibris i.d.R. Blüten nur im Schwirrflug anfliegen und klammern sich nur ausnahmsweise an größeren Blüten fest. Als Blütentypus erscheint bei von Kolibris bestäubten Vogelblumen besonders häufig der Röhren- oder Kelchtyp, wie er u.a. für Lippenblütengewächse (Lamiaceae), Braunwurzgewächse (Scrophulariaceae), Nachtschattengewächse (Solanaceae), Gesneriengewächse (Gesneriaceae) oder eine Reihe von Baum- und Straucharten sowie Epiphyten (z.B. einige Orchidaceae, Bromeliaceae) typisch ist. Die Staubblätter überragen häufig die Länge der Kronröhre, wobei der Pollen entweder auf den Schnabel, die Stirn bzw. den Hinterkopf oder das Kinn des Kolibris übertragen wird (Abb. 2).
Über die Entstehung der Nektarivorie bei Vögeln wie auch umgekehrt der Ornithophilie im Pflanzenreich gibt es zahlreiche, teils widersprüchliche Hypothesen, die selbst eine Deutung im evolutionären Konzept erschweren. In der modernen Forschung wird der früher häufig strapazierte Begriff der Koevolution zunehmend kritisch gesehen und die Pflanze-Vogel-Beziehung lediglich als Mutualismus gedeutet. Demzufolge dürfte es sich eher um ein stammesgeschichtlich gesehen relativ junges Phänomen handeln. Für diese Überlegung sprechen folgende Indizien: 1. Vogelblumen und ihre Bestäuber sind weltweit in den Tropen und Subtropen verbreitet. Ornithophilie ist zwar in vielen Pflanzenfamilien zu finden, jedoch nur bei jeweils wenigen Vertretern. Daher wird angenommen, daß dieses Phänomen mehrfach parallel bzw. unabhängig voneinander in verschiedenen Stammeslinien entstanden ist. 2. Nur wenige Vogelfamilien ernähren sich (neben Insekten, Früchten u.a.) überhaupt nektarivor (Abb. 1). Ausnahmslos handelt es sich dabei um relativ spezialisierte Vertreter mit entsprechenden anatomisch-morphologischen Konstruktionen (z.B. Schnabel-, Zungenform). Allerdings weisen nur die Kolibris eine extreme Spezialisierung hinsichtlich der Art der Nahrungsaufnahme und damit verbundener Anpassungen in Körperbau und Verhalten (z.B. Flügelkonstruktion - Schwirrflug) auf. 3. Trotz der zugrundeliegenden Spezialisierungen bei Vogel und Nahrungspflanze - als unmittelbare Voraussetzung für eine überhaupt erfolgreiche Ernährung bzw. Bestäubung - finden sich zumeist keine ausgeprägten "Exklusivbeziehungen", d.h. direkte Abhängigkeitsverhältnisse. Die Tatsache, daß einige Kolibriarten fast ausschließlich an bestimmte Nahrungspflanzen gebunden sind (Bsp.: Eutoxeres spp. - Heliconien), deutet nicht grundsätzlich eine parallele Entwicklung beider Taxagruppen an. Die sehr variable Schnabelform von Trochiliden (vgl. Weller 1999) stellt vorrangig die Bandbreite der Mikroevolution dar. Das Vorhandensein extremer morphologischer Merkmale bzw. Spezialisierungen - hierzu gehört ebenfalls die Veränderung der Körpergröße - muß selbstverständlich auch unter dem Aspekt der Konkurrenzvermeidung unter suboptimalen Bedingungen verstanden werden. Die innerhalb einer Gattung am weitesten verbreiteten und damit unter den Bedingungen adaptiver Radiationen erfolgreichsten Kolibris aus der Gruppe der Trochilinae (z.B. Amazilia-, Chlorostilbon-Arten, Florisuga mellivora) besitzen gerade bis schwach gebogene, mittellange Schnäbel (20-30 mm) und eine mittlere Körpergröße (8-12 cm).
4. Vielfach kann an Blüten mit relativ langen Kronblättern "Nektardiebstahl" beobachtet werden. Dabei stechen Arten mit kürzeren Schnäbeln Blütenkelche seitlich an, um an den Nektar zu gelangen; Bestäubung findet nicht statt. Von einigen Kolibriarten ist bekannt, daß sie Stechöffnungen nutzen, die von anderen Vögeln mit kräftigeren Schnäbeln (z.B. Coereba-, Diglossa-Arten) angelegt wurden. Ein derartiges Verhalten spricht selbstverständlich nicht für eine koevolutive Anpassung, sondern geschieht vermutlich entweder situationsbedingt oder gezielt und könnte teilweise erlernt worden sein. Im Rahmen des Evolutionsmodells werden insektenfressende Vögel als Vorfahren heutiger nektarivorer Gruppen angesehen (u.a. Faegri & van der Pijl 1966, Poley 1994). Insektennahrung ist auch bei Kolibris weit verbreitet (vor allem Schattenkolibris) und wird bei der Jungenaufzucht ausschließlich verwandt. Wie es zum Übergang von der einen in die andere Ernährungsform kam, ist umstritten. Die am häufigsten genannte Hypothese ist, daß zunächst Insekten aus Blüten erbeutet wurden und dabei allmählich Nektar als zusätzliche Nahrungsquelle entdeckt wurde. Eine vergleichbare Interpretationsmöglichkeit liefert die Vermutung, Vögel hätten sich zunächst primär von Wasser aus Blütenkelchen ernährt. Weniger plausibel erscheint, daß über die destruktive Aufnahme von Blütenteilen eine Gewöhnung an den Nektar erfolgte (vgl. Schnabelformen, Abb. 1) oder daß sich die Vorfahren nektarfressender Vögel zuerst von Baumsäften ernährten. Diese Ernährungsform ist zwar auch bei einigen Kolibriarten zu beobachten, erscheint jedoch lediglich als situationsbedingte Verhaltensweise. Vergleichbar der "illegalen" Nektaraufnahme aus beschädigten Blüten werden in diesem Fall beispielsweise von Spechten geschaffene Löcher genutzt, was nicht als mikroevolutive Anpassung zu verstehen ist. |
Die äußerst erfolgreiche Ausbreitung der Kolibris, die durch die Einnischung in der Ernährungsform begünstigt wurde, steht in engem Zusammenhang mit der geohistorischen Entwicklung Südamerikas. Das fast vollständige Fehlen von Fossilien - es sind lediglich einzelne Funde aus pleistozänen Schichten von karibischen Inseln bekannt - erschwert zweifellos die Beurteilung ihrer Stammes- und Artbildungsgeschichte. Dies ist aber kein Indiz für eine relativ junge Entstehung der Trochiliden, da aufgrund der besonderen geologischen Situation in Südamerika wie der Abtragungsrate in den Anden und der großräumigen Verfrachtung von Sedimenten im Amazonasbecken auch von anderen wichtigen Wirbeltiergruppen (u.a. Reptilien, Säuger) kaum fossile Nachweise bekannt sind. Erkenntnisse über Zentren und Mechanismen der Ausbreitung der Kolibris können daher vorrangig anhand von biogeographischen Untersuchungen, d.h. Analysen zur Verbreitung und morphologischen Variation gewonnen werden. Vergleichende Studien zu molekularbiologischen Ähnlichkeiten, z.B. DNA-Hybridisierungsexperimente (Sibley & Ahlquist 1990, Bleiweiss et. al. 1994, 1997), sollten auf der Grundlage biogeographisch-morphologischer Befunde erfolgen (Weller 1998).
Als Ursache (bzw. Auslöser) vielfältiger Artbildungsprozesse und damit der rezent beobachteten Vielfalt der neotropischen Tier- und Pflanzenwelt können Änderungen in der Geomorphologie des südamerikanischen Kontinents angenommen werden. In der frühen biogeographischen Literatur wurden die der Entstehung der Anden zugrundeliegenden Hebungsvorgänge als entscheidender Auslöser für Artbildungsprozesse bewertet (Chapman 1917, 1926). Diese Vorgänge begannen bereits mit dem Ende des Tertiärs am Übergang zum Quartär und sind bis heute nicht abgeschlossen. Für den am besten untersuchten Teil der Kordilleren, die Nordanden Kolumbiens und Ecuadors und angrenzende Gebirgszüge Mittelamerikas (vgl. Troll 1930), wird als Höhepunkt der orogenetischen Hebungen (= Entstehung von Gebirgsstufen über 2000 m) das frühe Pleistozän (vgl. Tab. 1), im Fall der Ostanden (z.B. Mérida-Region, Venezuela) das mittlere bis späte Pliozän angesehen (Simpson 1979). Aufgrund von Studien über Pollenablagerungen in aufeinanderfolgenden Sedimenten ausgewählter Gebirgs- und Tieflandgebiete Südamerikas (z.B. Bogotá-Region, Kolumbien; Manaus, Brasilien) wird postuliert, daß Vegetationsfluktuationen stattgefunden haben, die auf die Temperaturschwankungen im Pleistozän zurückzuführen sind und vom Wechsel von Eiszeiten und Zwischeneiszeiten beeinflußt wurden (van der Hammen 1974, van der Hammen & Gonzales 1960, 1964, Haffer 1967). Im amazonischen Tiefland kam es während glazialer Schübe in kühl-trockenen Perioden zur Ausbreitung von Savannengebieten, was die erhöhte Ablagerung von Gräserpollen zur Folge hatte. Umgekehrt konnten sich Wälder unter warm-feuchten Bedingungen ausbreiten (überwiegende Ablagerung von Pollen von Baumarten). In den Anden wurde der vertikale Temperaturgradient im Wechsel kälterer und wärmerer Phasen entweder erhöht oder verringert, was Auswirkungen auf die Höhenverbreitung der Vegetation, insbesondere der Bergregenwälder, hatte. Ihre Vertikalverteilung dürfte für die Einnischung bestimmter Vogelgruppen demnach weitaus entscheidender für Artbildungsprozesse gewesen sein als rein geologische Vorgänge, wie u.a. Graves (1985) am Beispiel der Diglossa-Arten (Blütenstecher) nachwies.
Die Analyse der Verbreitung tropischer Pflanzenfamilien des Amazonasbeckens (Prance 1973) sowie die bereits erwähnten Pollenfunde (van der Hammen 1974) geben Hinweise auf Verbreitung und Größe ehemaliger Waldinseln. Demnach werden im nördlichen Südamerika neun Haupt- und eine Reihe weiterer Nebenrefugien vermutet. Geologisch gesehen befinden sich die meisten im Bereich der Anden bzw. des Guyana-Schildes (Haffer 1969). In den Refugien bildeten überlebende Populationen die Ausgangsbasis für Radiationen. Relativ häufig differenzierten sich Refugialpopulationen auf Art- oder Unterartebene, bevor sie sich unter günstigeren ökologischen Bedingungen geographisch weiter ausbreiten konnten. Falls die Zeiträume der Isolation ausreichend lang andauerten, um eine ökologische bzw. morphologische Differenzierung der Taxa zu ermöglichen, war eine Hybridisierung infolge von späteren erneuten Überlappungen der Verbreitungsgebiete (Sekundärkontakte) mit ehemaligen Vorläuferpopulationen nicht mehr möglich und somit der Prozeß der allopatrischen Artbildung abgeschlossen (Übersicht in Haffer 1979). Geomorphologische Befunde hinsichtlich der Sedimentationsraten in andinen Tälern (Simpson 1979) lassen darauf schließen, daß in Trockenzeiten vermehrt Erosionsprozesse aufgrund teilweiser Entwaldung auftraten. In feuchteren Perioden wurden die Sedimente dann ausgewaschen. In der Pantepui-Region SO-Venezuelas kam es umgekehrt zur Entwaldung von Flanken der Tafelberge, die als mächtige Gesteinsschichten nahezu senkrecht abgetragen wurden. Beide Prozesse hatten signifikante Veränderungen der Topographie betroffener Regionen zur Folge und wirkten vermutlich als Isolationsfaktoren artbildungsfördernd (Steyermark 1986). Aufgrund von Studien zu Endemismusraten (Mayr & Phelps 1967) wird angenommen, daß die Vorläufer vieler heutiger Vogelarten die Tafelberge erst in jüngerer Zeit (z.B. Pleistozän) besiedelten. Endemische Kolibriarten lassen sich wahrscheinlich überwiegend von andinen Vertretern ableiten und benutzten flußbegleitende Wälder als Einwanderungsbrücken (Weller im Druck). Generell bietet das Refugialmodell eine befriedigende Erklärung für die heutige Verbreitung vieler neotropischer Kolibriarten. Biogeographische Analysen tieflandbewohnender (z.B. Agyrtria, Amazilia, Polyerata, Weller 1998) als auch andiner Trochilidengattungen (z.B. Aglaiocercus, Schuchmann & Duffner 1993; Chalcostigma, Schuchmann & Heindl 1997) lassen häufig Kongruenzen von Entstehungs- oder Verbreitungsgebieten mit postulierten Wald- oder Offenlandrefugien erkennen. Neben der Theorie der Differenzierung von Arten oder Unterarten durch Isolation in Refugien werden weitere Modelle diskutiert, die Unterschiede in regionalen bzw. kleinräumigen Umweltbedingungen als Auslöser für Artbildungsprozesse betonen. Alternative DifferenzierungsmodelleDie Refugientheorie ist nicht unumstritten (Endler 1982, Connor 1986, Salo et al. 1986), obwohl die postulierten Endemismuszentren unterschiedlicher Tiergruppen im Wesentlichen übereinstimmen (vgl. Brown & Mielke 1972, Heliconia-Falter; Vanzolini & Williams 1970, Anolis-Echsen). Alternativ zur Annahme von Artbildung in plio-pleistozänen Refugien (entsprechend dem peripheral isolates model, vgl. Mayr 1967) wies Fjeldså (1995) auf die Möglichkeit unabhängiger Radiationen von Reliktpopulationen in ökologisch extrem stabilen, im gesamten Pleistozän und unabhängig von Refugien existierenden Gebirgsarealen hin ("stable area" theory). Artbildung unter diesen Voraussetzungen bezeichnet man als Zentrifugalspeziation (Frey 1993, basierend auf Brown 1957). Sie steht im Gegensatz zu Vikarianzeffekten, d.h. Artbildungsprozessen, die durch ökologische oder geographische Barrieren hervorgerufen worden sind. Beispielgebend betonte Fjeldså (1990, 1995) die Bedeutung andiner Feuchtwälder (Peru, Chile, Bolivien) - die häufig von Polylepis (Rosengewächse) gebildet werden - als Endemismusgebiete (Entstehungs- bzw. Verbreitungszentren) für gebirgsbewohnende Vogelarten. Auch Kolibrivertreter gehören zu den Bewohnern solcher Gebirgswälder und entstanden möglicherweise in solchen "stable areas", worauf u.a. Heindl & Schuchmann (1998) bei der Gattung Metallura hinwiesen. Demnach spricht vieles dafür, daß sich diese Gruppe wie viele andere Vogeltaxa erst im späten Pleistozän diversifizierte (vgl. Klicka & Zink 1997) und nicht bereits im frühen Pleistozän vor ca. 2-3 Millionen Jahren (nach Garcia-Moreno et al. 1999). Diese Hypothese wird durch Befunde gestützt, die starke klimatische Oszillationen während der letzten 800.000 Jahre im Bereich der Nordanden vermuten lassen, und zwar stärkere in 100.000-Jahres-Zyklen (entsprechend den Glazial-Interglazial-Perioden) und schwächere, aber häufigere (19.000-23.000 J.) während des gesamten Pleistozäns (Hooghiemstra et al. 1993). Die Beobachtung, daß Verbreitungsgebiete von Tier- und Pflanzenarten in Amazonien häufig mit dem Verlauf von Flußsystemen korrelieren, führte einige Autoren zur Vermutung, daß breite Flußläufe potentielle Barrieren des Genflusses zwischen Populationen darstellen und deshalb als Artbildungsfaktoren wirken. Für die Vogelwelt wiesen u.a. Sick (1967) und Haffer & Fitzpatrick (1985) auf den Einfluß von Flüssen als Ausbreitungsschranken insbesondere auf unterwuchsbewohnende, relativ standorttreue Arten hin. Für Vogelarten mit höherem Ausbreitungspotential wirken sie im allgemeinen nicht als artbildungsfördernde Barrieren. Dies konnte auch für einige in Amazonien weitverbreitete Kolibris der Gattungen Agyrtria und Polyerata nachgewiesen werden (Weller 1998). In beiden Gruppen kommen Vertreter derselben Art beiderseits des Amazonas vor und unterscheiden sich entweder nur auf Unterartniveau (Agyrtria versicolor) oder nicht signifikant und zeigen mehr oder minder deutliche Hybridmerkmale (Polyerata fimbriata). | |||||||||||||||||||
Entsprechend der Progressionsregel Hennigs (1950) lassen sich plesiomorphe, d.h. ursprüngliche Merkmale vor allem bei stammesgeschichtlich älteren Vertretern nachweisen, während jüngere Vertreter einer Tiergruppe verstärkt durch apomorphe, d.h. "fortschrittliche" Kennzeichen charakterisiert sind. Bei den Trochilidae wird die Unterfamilie der Schattenkolibris (Phaethornithinae) allgemein als relativ ursprünglich betrachtet, während die Eigentlichen Kolibris (Trochilinae) als stärker abgeleitet erscheinen (Johnsgard 1983). Für diese Theorie sprechen folgende Merkmale und Lebensweisen: 1. Im Gefieder zeigen Schattenkolibris als überwiegend waldbewohnende Arten düstere, nichtmetallische Farben, während die meisten anderen Vertreter Farbglanzeffekte aufweisen, möglicherweise eine Anpassung an die vielfältigen Lebenräume dieser Unterfamilie. 2. Alle Schattenkolibris zeigen kaum farbliche Unterschiede der Geschlechter, während die Mehrzahl der Trochilinae geschlechtsdimorph ist, d.h. männliche und weibliche Individuen einer Art sind verschieden gefärbt. Männchen besitzen häufig auffällige gefärbte Schmuckfedern oder Körperanhänge, was auf sexuelle Selektion im Rahmen eines stärker ausgeprägten Balzverhaltens zurückgeführt werden kann (z.B. Schauflüge). 3. In der Nahrungswahl bevorzugen Schattenkolibris einen höheren Anteil Insekten, die Eigentlichen Kolibris eher Blütennektar. Insekten werden als ursprünglichere Nahrungsform nektarivorer Vögel angesehen (vgl. Theorien zur Entstehung der Nektarivorie). 4. Als biogeographisches Zentrum der Familie der Kolibris wird das nordöstliche Südamerika postuliert, da Formen des westlichen Südamerikas und Mittelamerikas mehr abgeleitete Merkmale zeigen. Obwohl das gesamte Verbreitungsgebiet der Schattenkolibris sowohl weite Bereiche Südamerikas als auch Mittelamerikas umfaßt, treten sie vor allem in Brasilien in zahlreichen Arten auf und haben dort offenbar ihren Ursprung. Hierbei ist anzumerken, daß nur wenige, allerdings relativ plesiomorph (ursprünglich) erscheinende Vertreter der Trochilinae wahrscheinlich ebenfalls in dieser Region entstanden sind (z.B. Gattungen Agyrtria, Polyerata; Weller 1998). Andere Merkmale und Eigenschaften weisen aber auch auf Spezialisierungen bei Schattenkolibris hin, die eher als apomorphe Charakteristika einzustufen sind. Im Körperbau fällt der relativ lange, gebogene Schnabel auf, eine spezielle Anpassung an Blüten mit gekrümmter Kronröhre (z.B. Bananengewächse, Heliconiaceae). Demgegenüber besitzen die meisten trochilinen Kolibris kürzere und gerade bis schwach gebogene Schnäbel, erscheinen also weniger spezialisiert. Das könnte andeuten, daß das Merkmal Schnabelform von einfachen (geraden) zu komplexeren (gekrümmten) Schnäbeln modifiziert wurde. Eine weitere morphologische Besonderheit stellt der abgestufte, teilweise kontrastierte Schwanz der Schattenkolibris dar. Er dürfte eher Bedeutung im Revier- und Balzverhalten besitzen, während der zumeist kurze, leicht bis mäßig gegabelte Schwanz der Eigentlichen Kolibris besser den grundlegenden Anpassungen und Strategien des Nahrungserwerbs, des Balz- und Brutverhaltens und auch der Feindabwehr (u.a. erhöhte Manövrierfähigkeit; verminderte Abstoßung während des Brütens) Rechnung trägt. Als weiteres Indiz für eine Spezialisierung der Schattenkolibris kann die eigentümliche Nestform gelten (vgl. Weller 1999). Die an Pflanzenblättern aufgehängte Konstruktion ist wahrscheinlich nur eine Modifizierung des Napfnestes, des bei den Eigentlichen Kolibris und den meisten anderen Vögelgruppen am häufigsten verbreiteten Nesttyps. Intermediäre Merkmale im Nestbau und in der Morphologie der Zahnschnabelkolibris (Androdon, Doryfera) lassen diese Artengruppe als Übergangsform beider Unterfamilien erscheinen (Schuchmann 1995). Die vorangehenden Ausführungen verdeutlichen, daß die Hypothesen über die Stammesgeschichte der Kolibris zu wenig gesichert sind, um definitive Schlußfolgerungen über den Ursprung der wichtigsten Artengruppen und ihrer Verwandtschaftsverhältnisse zu ziehen. Eine Deutung ihrer Herkunft und Evolution im Grundtypmodell (Definition bei Junker & Scherer 1998) erscheint ebenfalls nicht unproblematisch, zumindest unter Berücksichtigung des Kriteriums der Kreuzbarkeit von Gattungen verschiedener Unterfamilien. Kreuzungen zwischen unterschiedlichen Kolibriarten oder -gattungen kommen in der Natur offenbar nicht selten vor und haben häufig zu taxonomischen Fehlschlüssen hinsichtlich der Gültigkeit von Arten geführt (Hinkelmann et al. 1991). Da intermediäre Merkmale in Körperbau und Gefiederfärbung als Resultat von Kreuzungen in der Vergangenheit häufig übersehen worden sind, wurde eine Reihe von Arten beschrieben, die sich bei genauerer Überprüfung als Hybriden erweisen. Insbesondere sind solche Artbeschreibungen kritisch zu betrachten, die nur auf der Untersuchung eines einzelnen Belegexemplares beruhen. Ein prominentes Beispiel dafür stellt der Fall der sog. Táchira-Amazilie (Amazilia distans, Wetmore & Phelps 1956) aus Venezuela dar, die kürzlich als Gattungshybrid Amazilia × Hylocharis identifiziert werden konnte (Weller & Schuchmann 1997, Graves 1998). Hybriden sind bislang ausschließlich aus der Unterfamilie der Eigentlichen Kolibris bekannt geworden. Insgesamt wurde Hybridisierung zwischen mehr als 30 Gattungen nachgewiesen (u.a. Hartert 1900, Berlioz 1927, 1929, 1938, 1964, Grantsau 1988, Graves 1996, 1997, eigene Studien). Zahlreiche Beispiele für direkte und indirekte Kreuzbarkeit liefern insbesondere die südamerikanischen artenreichen Gattungen Amazilia, Polyerata, Hylocharis, Thalurania und Chlorostilbon sowie die nordamerikanischen Arten der Gattungen Selasphorus, Stellula und Archilochus bzw. Calypte (Short & Phillips 1966). Zweifelsohne deutet das gehäufte Auftreten von Hybridexemplaren auf enge phylogenetische Beziehungen der beteiligten Taxa hin, worauf Mayr & Short (1970) hinwiesen. So wird z.B. für die in Nordamerika endemischen Kolibrivertreter Monophylie, d.h. eine gemeinsame phylogenetische Abstammung von süd- und mittelamerikanischen Vorfahren angenommen (Johnsgard 1983). Unterschiede in wesentlichen morphologischen und verhaltensbiologischen Merkmalen (z.B. Bioakustik, Brutbiologie) sind vermutlich die Ursache für fehlende Kreuzungsnachweise zwischen den Eigentlichen Kolibris und den Schattenkolibris (einschließlich der Zahnschnabelkolibris). Beide Unterfamilien werden als monophyletisch angesehen. Wahrscheinlich haben sich ihre Evolutionslinien in einem frühen Stadium der Stammesgeschichte der Kolibris getrennt und unabhängig voneinander weiterentwickelt. Allerdings sind alle Vertreter der Kolibris durch den gemeinsamen Besitz einer Kombination einzigartiger Merkmale von anderen, auch habituell ähnlich erscheinenden Vogelfamilien deutlich abgrenzbar. Deshalb erfüllen Kolibris durchaus die Anforderungen an einen Grundtyp. Die teilweise starken Unterschiede in Morphologie und Verhalten sind im Rahmen der Variabilität von Grundtypen und der Polyvalenz von Stammformen interpretierbar (Junker & Scherer 1998). Vor diesem Hintergrund erscheint die heutige Formenvielfalt der Trochiliden deshalb als Resultat vielfacher paralleler und aufeinanderfolgender Artbildungsprozesse unter den sich rasch verändernden geologischen und klimatischen Bedingungen Südamerikas. Dank: Mein herzlicher Dank gilt der SG Wort und Wissen e.V. für ihre finanzielle Unterstützung meiner Promotion, in deren Verlauf ich mich in die vorangehende Thematik einarbeiten konnte. Für logistische Hilfe und Betreuung sowie die Überlassung von Arbeitsmöglichkeiten danke ich dem Zoologischen Forschungsinstitut und Museum A. Koenig, Sektion Ornithologie, Bonn, sowie PD Dr. K.-L. Schuchmann für eine kritische Diskussion meiner Ergebnisse und o.g. Fragestellungen. |

|