|
|
|
|
|
Protarchaeopteryx
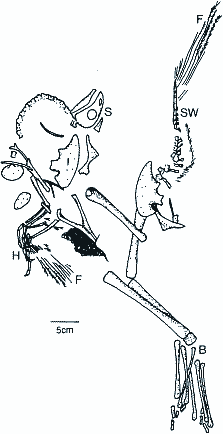
CaudipteryxGroße Ähnlichkeit zu Protarchaeopteryx zeigt Caudipteryx (deutsch "Schwanzfeder"), ebenfalls ein Vertreter der Theropoda (vgl. Tab. 1). Caudipteryx ist mit einer Länge von 70 cm ein relativ kleiner Vertreter dieser Gruppe. Er hat nur vier Zähne. Das Schwanzskelett entspricht in seinen Proportionen etwa dem des Archaeopteryx, Arm- und Handskelett sind allerdings ziemlich kurz (Abb. 2). Auch Caudipteryx zeichnet sich durch den Besitz von Federn aus, hier fossil erhalten am zweiten Mittelhandknochen und Finger sowie am Ende der Schwanzwirbelsäule. |
Für diese Annahme sprechen auch die Proportionen ihres Arm- und Handskeletts; sie waren für einen aktiven Flug zu schwach. Bei Caudipteryx hat der Arm nur 60% von der Länge seines Beins, im Fall von Protarchaeopteryx ist die Armlänge nur geringfügig größer. Archaeopteryx dagegen hat etwa gleichlange Arme und Beine. Die Federn zeigen etwa die gleiche Tendenz, d.h. die Schwungfedern von Archaeopteryx sind um einiges länger. Der Erhaltungszustand der Fossilien läßt über den mikroskopischen Feinbau der Saurier-Federn noch keine endgültigen Aussagen zu. Protarchaeopteryx und Caudipteryx sind als Vorfahren von Archaeopteryx auch dann fragwürdig, wenn man ihre symmetrischen Federn als Vorstufe der asymmetrischen Archaeopteryx-Feder deuten würde, denn das Armskelett von Protarchaeopteryx und Caudipteryx war deutlich zu kurz, um es zum Fliegen einsetzen zu können. Wozu hatten sie also komplexe Federn entwickelt? Ein sekundärer Flugverlust erscheint naheliegender. Damit würden sie nur einen blind endenden Seitenast im Stammbaum der Vögel besetzen.
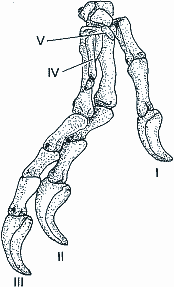
Auch aus anderen Gründen sind Zweifel an dem Schluß angebracht, die bisher gefundenen befiederten Saurier seien die Vorfahren der modernen Vögel. So macht Hinchliffe (1997) auf die Schwierigkeit aufmerksam, aus der im Vergleich zur Gesamtkörpergröße sehr kleinen Vorderextremität der Theropoden einen Vogelflügel abzuleiten. Burke und Feduccia (1997) haben zudem nachgewiesen, daß die Finger der Dinosaurier nicht mit den Fingern einer Vogelhand gleichgesetzt werden können. So zeigt zum Beispiel der zweibeinige Theropode Herrerasaurus, daß die gewöhnlich dreifingerige Theropodenhand die Finger I (Daumen), II (Zeigefinger) und III (Mittelfinger) ausgebildet hat, während die Finger IV und V reduziert (bei Herrerasaurus noch als kleine Stummel zu erkennen) sind (Abb. 3). Dagegen sind bei der Vogelhand stets (von den Autoren in zahlreichen embryologischen Studien nachgewiesen) die Finger II, III und IV ausgebildet, während hier I und V im Laufe der Embryonalentwicklung reduziert werden.
Die zeitliche Einordnung der befiederten Dinosaurier schließt Protarchaeopteryx und Caudipteryx ebenfalls als Vorfahren des Archaeopteryx aus. Das Alter der Schichten, in denen sie gefunden wurden, wird mit 120 bis 135 Millionen Jahren angegeben (eine Gruppe kanadischer Geologen legt sich auf 121 bis 123 Millionen Jahre fest); den Schichten der Solnhofener Plattenkalke, aus denen die Archaeopteryx-Fossilien geborgen wurden, schreibt man dagegen ein Alter von 150 Millionen Jahren zu. Somit kommen die jüngeren befiederten Dinosaurier nicht als Vorfahren des älteren Archaeopteryx in Frage. SinosauropteryxEin weiterer Fund eines theropoden Raubdinosauriers aus der Yixian-Formation verdient Erwähnung. Sinosauropteryx - so der Name des in mehreren Exemplaren gefundenen Fossils - hat eine gewisse Ähnlichkeit mit Compsognathus, dem aus Deutschland (Solnhofen) bekannten theropoden Dinosaurier (vgl. Tab. 1) und mutmaßlichen Zeitgenossen von Archaeopteryx. Sinosauropteryx war mit 65 cm Länge ebenfalls ein recht kleiner Saurier. Er lief aufrecht auf den Hinterbeinen, hatte kurze Arme und an diesen drei Finger mit Krallen, des weiteren einen sehr langen Wirbelschwanz. Das besondere Interesse der Paläontologen galt zunächst faserigen Strukturen entlang der Wirbelsäule von Sinosauropteryx, die von einigen Forschern als primitive Federn interpretiert wurden (Abb. 4). Mittlerweile ist man in Fachkreisen aber ziemlich sicher, daß diese zunächst als "Vor-Federn" angesehenen Fasern wohl eher Kollagenfasern aus dem Bindegewebe unter der Haut von Sinosauropteryx sind (Gibbons 1997). Ähnliche Strukturen gibt es beispielsweise auch im Bindegewebe von Seeschlangen (unter der Haut im Schwanzbereich). Dennoch bleiben einige Paläontologen bei der "Vor-Feder-Theorie". Weitere Untersuchungen, vor allem auch die Beantwortung der Frage, ob es sich hier um Keratin (dem Stoff, aus dem die Federn sind) oder Kollagen handelt, können hier vielleicht Klarheit schaffen. |
Inzwischen sorgt ein weiteres Detail aus sehr gut erhaltenen Exemplaren von Sinosauropteryx für heiße Diskussionen. Im Hinterleib einiger Fossilien zeichnet sich ein mehr als 5 cm langer ovaler dunkler Fleck ab, der im Brustbereich mit einer Wölbung endet. Ruben et al. (1997) interpretieren diese Bildung als Zwerchfell und vergleichen sie mit dem Zwerchfell bei Krokodilen. Rezente Krokodile besitzen tatsächlich eine zwerchfellähnliche Membran aus einer dicken Lage von Bindegewebe, die Brustraum und Bauchraum voneinander trennt. Diese Membran wirkt zwar wie ein Zwerchfell und sorgt für die Ventilation der Lunge, enthält aber keine Muskelfasern (wie z.B. bei Säugetieren). Die Atembewegung der Krokodile kommt zustande durch einen Rückziehmuskel an der Leber. Dabei wirkt die Leber mit ihrer Lage zwischen Membran und Rückziehmuskel wie ein Pumpenkolben. Eine anatomische Besonderheit der Krokodile besteht somit darin, daß die Leber kuppelförmig in den Brustraum hineinragt. Die genannten Autoren sehen bei Sinosauropteryx einen entsprechenden Befund. Der dunkle Fleck im Bauchraum des kleinen Kreide-Sauriers wird von ihnen als Leber bzw. als fossiler Überrest der Organe des Bauchraums interpretiert, der kuppelförmige Abschluß zum Brustraum als Zwerchfell (Abb. 5). Das würde bedeuten, daß die Atmung bei Sinosauropteryx entsprechend der der rezenten Krokodile funktionierte. Jüngst veröffentlichte Ergebnisse von Weichteilstudien des Theropoden Scipionyx bestätigen diese Interpretation (Ruben et al. 1999). |
|
Hieraus ergibt sich die Frage nach einer möglichen phylogenetischen Ableitung der Vögel. Wenn Sinosauropteryx und damit vielleicht alle Dinosaurier in der Anatomie der Atemorgane so hochgradige Übereinstimmungen mit den Krokodilen zeigen, dann ist es undenkbar, daß sich Vögel aus Dinosauriern entwickelt haben. Die Vogellunge besitzt nämlich eine völlig andere Konstruktion, die nach einem ganz anderen Prinzip arbeitet. Zunächst besitzen Vögel kein Zwerchfell. Im Gegensatz zu den Krokodilen sind bei ihnen die Lungenflügel volumenkonstant und starr. Beim Einatmen vergrößert sich der Brustraum der Vögel durch seitliches Auseinanderweichen der Rippen. Die eigentliche Ventilation besorgen die mit der Lunge verbundenen Luftsäcke, die diese Bewegung mitmachen. Ein weiterer wesentlicher Unterschied besteht im Fehlen von blind endenden Alveolen (Lungenbläschen). Die Vogellunge besitzt stattdessen ein durchlaufendes Röhrensystem. Die Luft strömt hier in einer Art Einbahnstraße durch die Lungenröhren in die Luftsäcke und dann durch andere Röhren wieder zurück (Abb. 6). Auf diese Weise ist sowohl beim Einatmen als auch beim Ausatmen Gasaustausch in blind endenden Luftkapillaren (die an Blutkapillaren anliegen) möglich. Durch dieses Wirksystem ist die Vogellunge das leistungsfähigste Atemorgan aller Wirbeltiere. |
|
Krokodile hingegen ventilieren ihre Lunge nach dem Blasebalgprinzip und besitzen Alveolen (Lungenbläschen, die von einem Netz von Blutkapillaren umgeben sind, an denen der Gasaustausch stattfindet). Ein Übergang von der einen zur anderen Konstruktion ist kaum denkbar. Des weiteren würde eine zweigeteilte Leibeshöhle, wie Krokodile sie besitzen, die Anlage von Luftsäcken verhindern. Wird die Trennung von Bauchraum und Brustraum jedoch aufgehoben, dann funktioniert das Zwerchfell nicht mehr. Eine funktionsfähige Konstruktion, die zwischen dem Prinzip "Blasebalglunge mit Alveolen" und dem Prinzip "Röhrenlunge mit Luftkapillaren" vermittelt, ist unvorstellbar. Ein weiteres gravierendes Problem ergibt sich bei der Embryonalentwicklung. Frisch geschlüpfte Krokodile füllen ihre Lunge mit dem ersten Atemzug, während junge Vögel schon Tage vor dem Schlüpfen im Ei atmen. Diese Atmung vor dem Schlüpfen ist notwendig für die Entwicklung der Röhrenlunge, die nicht plötzlich aufgeblasen werden kann. Sie setzt aber auch eine völlig andere Beschaffenheit des Vogeleis im Vergleich zum Reptilienei voraus, denn der Vogelembryo benötigt einen Luftvorrat im Ei sowie eine poröse Eierschale. Eine phylogenetische Ableitung der Embryonalentwicklung der Vögel von der der Krokodile ist also nicht so einfach möglich. Es gibt noch weitere Gründe, die gegen die Krokodile als Vorfahren der Vögel sprechen. Zum Beispiel haben Vögel ein Gabelbein. Dieses leitet man von zwei verschmolzenen Schlüsselbeinen ab. Krokodile jedoch haben kein Schlüsselbein und kommen daher als Urahnen der Vögel nicht in Frage. Die Anatomie des Beckengürtels der Krokodile schließt ebenfalls eine nähere Verwandtschaft mit den Vögeln aus. Es muß erwähnt werden, daß es Vorbehalte und Skepsis gegenüber der Ansicht gibt, die Dinosaurier hätten eine Krokodillunge besessen. Die schwarzen Flecken in der Leibeshöhle von Sinosauropteryx müssen schließlich nicht zwingend als Reste von Leber und Zwerchfell interpretiert werden. Die von Ruben et al. (1999) jüngst präsentierten Befunde weisen aber erneut auf eine starke Ähnlichkeit der Theropodenlunge und der Krokodillunge hin (vgl. auch Wuethrich 1999). Man darf die weitere Entwicklung dieser Diskussion und vielleicht auch weitere Funde aufschlußreicher Fossilien mit Spannung erwarten. Für den Moment ergeben sich einmal mehr aus den äußerst interessanten neuen Fossilfunden, von dem man Rückschlüsse auf den Verlauf der Stammesgeschichte erwartet (hat), mehr Fragen als Antworten. |
In diesem Falle nicht aus China, sondern von der Insel Madagaskar wird ein weiterer interessanter Fund eines fossilen Vogels gemeldet. Rahona ostromi wird ein Alter von 60 bis 70 Millionen Jahren zugeschrieben. Er hatte befiederte Schwingen wie ein moderner Vogel, andererseits aber eine lange, knöcherne Schwanzwirbelsäule und eine stark ausgeprägte sichelförmige Kralle an der zweiten Zehe. Letztere erinnert sehr stark an eine entsprechende Bildung bei dem theropoden Raubdinosaurier Velociraptor. Die fossilen Überreste des etwa rabengroßen Rahona aus der Oberkreide Nordwest-Madagaskars werden von Forster et al. (1998) als Mosaik primitiver Merkmale - übereinstimmend mit dem Bauplan theropoder Dinosaurier - und abgeleiteter Vogel-Merkmale beschrieben. Typisch für einen Theropoden sind z.B. das Beinskelett und der Bau der Wirbel, vogeltypisch dagegen u.a. die opponierte erste Zehe (nach hinten gerichtete Zehe zum Umgreifen eines Astes) und die hohlen Knochen. In einigen Merkmalen ist Rahona sogar "moderner" als Archaeopteryx (beispielsweise durch den Besitz verschmolzener Sakralwirbel). Nach geochronologischen Befunden ist Rahona jedoch ca. 80 Millionen Jahre jünger als Archaeopteryx. Diese Unstimmigkeit veranlaßte einige Forscher dazu, an der Identität des Fossils zu zweifeln. So erwähnt Gibbons (1998) den Verdacht, daß es sich bei Rahona in Wirklichkeit um zwei Fossilien (also einen Vogel und einen Saurier) handeln könnte, die nach vorherigem Zerfall gemeinsam abgelagert wurden. Bleibt man jedoch bei der Interpretation, daß es sich bei Rahona nur um ein einziges Fossil handelt, dann läßt sich für diese Mosaikform mit sehr primitiven und stark abgeleiteten Merkmalen ein Platz im Stammbaum der Vögel nur dann finden, wenn man sie auf einen blind endenden Seitenzweig setzt und erhebliche Konvergenzen annimmt. |
Nach dem derzeitigen Stand der Forschung sind die neu entdeckten Gattungen Protarchaeopteryx und Caudipteryx zweifelsfrei theropode Saurier, die Federn (mit bisher unbekannter Feinstruktur) trugen. Ihre symmetrischen Federn waren aber ebenso wie ihr Arm- und Handskelett für einen aktiven Flug ungeeignet und sind in Analogie zu heutigen Verhältnissen am ehesten als sekundär zu deuten. Als Vorstufen zu flugtauglichen Federn kommen sie kaum in Betracht, weil die geschilderten anatomischen Verhältnisse dazu nicht passen. Somit scheiden sie als Vorfahren des Archaeopteryx und der "modernen" Vögel aus. Die Datierung der entsprechenden Schichten spricht ebenfalls gegen eine Abstammung des Archaeopteryx und der modernen Vögel von Protarchaeopteryx oder Caudipteryx. Wertete man sie dennoch als Vorfahren der Vögel, müßte Archaeopteryx als parallel entstandener Seitenzweig betrachtet werden. Federn wären demnach mindestens zweimal unabhängig entstanden - evolutionstheoretisch sehr problematisch. Die Funktion der Befiederung bei Protarchaeopteryx und Caudipteryx muß spekulativ bleiben (Isolation, Tarnung, Balz?); unklar ist, weshalb Federn so komplex gebaut sind, wenn sie ursprünglich nicht zum Fliegen benötigt worden wären. Anatomie und Embryologie des Handskeletts schließlich sprechen generell gegen eine Ableitung der Vögel von den theropoden Sauriern. Eine stammesgeschichtliche Beziehung zwischen Archaeopteryx und den befiederten Sauriern ist nur möglich, wenn man annimmt, daß beide von bereits flugfähigen, befiederten Sauriern abstammten, die im Falle von Protarchaeopteryx und Caudipteryx ihre Flugfähigkeit wieder einbüßten, im Falle von Archaeopteryx dagegen beibehielten. Da auch Archaeopteryx schon seit einiger Zeit aufgrund spezieller Merkmale nicht als Vorfahre der modernen Vögel diskutiert wird (vgl. Junker & Scherer 1998), ist evolutionstheoretisch die nächstliegende Schlußfolgerung, sowohl die (bisher gefundenen) befiederten Saurier als auch Archaeopteryx auf einen blind endenden Seitenzweig des Vogelstammbaums zu plazieren (Abb. 7). |
|
|
Die Überlieferung früher Vögel und befiederter Saurier paßt somit am besten zur Annahme einer polyphyletischen Entstehung verschiedener Konstruktions- und Mosaikformen mit unterschiedlichen Kombinationen von Vogel- und Sauriermerkmalen. Diese Sichtweise drängt sich auch bei der Deutung der Vielfalt kreidezeitlicher Vogelgruppen auf, die nicht in widerspruchsfreie Stammbaumabfolgen eingeordnet werden können (vgl. Fehrer & Zimbelmann 1998). Auch die Vermutung, Sinosauropteryx (und vielleicht alle Dinosaurier) hätten eine Lunge, die wie eine Krokodillunge funktionierte, spricht eher für eine Interpretation im Sinne mosaikartiger, voneinander unabhängiger Formen als für einen evolutiv gangbaren Weg von Reptilien zu Vögeln. Die Entwicklung der Vogellunge aus der Krokodillunge ist aus anatomischen, funktionalen und embryologischen Gründen jedenfalls undenkbar. Das Fossil Rahona schließlich ist eine extreme Mosaikform, deren Kombination von Merkmalen theropoder Saurier mit denen moderner Vögel wohl nur mit dem merkwürdigen Bauplan des Schnabeltiers vergleichbar ist, welches Säugerhaare, den Besitz von Michdrüsen, eierlegende Fortpflanzungsweise, einen in mancher Hinsicht entenähnlichen Schnabel mit speziellen Merkmalen (z. B. einem Ruderschwanz) kombiniert. Die Euphorie der Tagespresse, wonach mit den neuen Dinosaurierfunden der Übergang Reptil (Dinosaurier) Die Überlieferung früher Vögel und befiederter Saurier paßt am besten zur Annahme einer polyphyletischen Entstehung verschiedener Mosaikformen. |
Literatur
|

|