|
Mikroorganismen als Energiekünstler
Der Energiehaushalt anaerober Mikroorganismen ist weder primitiv noch ineffizient
von Boris Schmidtgall
Studium Integrale Journal
26. Jahrgang / Heft 1 - April 2019
Seite 27 - 34
|
 |
Zusammenfassung: Über 30 Jahre lang waren Forscher der Ansicht, dass der Energiestoffwechsel anaerober Mikroorganismen aufgrund des Fehlens sogenannter Bifurkations-Enzyme deutlich weniger effizient sei als derjenige höherer Organismen – eine Auffassung, die aus der Perspektive der Darwin’schen Evolutionslehre durchaus plausibel erscheint. Doch jüngere Untersuchungen an verschiedenen Anaerobiern zeigen, dass auch diese vermeintlich primitiven Lebewesen mit erstaunlich anspruchsvollen biochemischen Funktionseinheiten ausgestattet sind.
|
|
|
Energie ist kostbar
Für jede Form der Bewegung und Thermoregulation (Wärmehaushalt) bedarf es der Energie. In diesem Zusammenhang wird oft allgemein vom „Energiehaushalt“ gesprochen. Ob Unternehmen, Maschinen, Häuser oder auch Lebewesen – alle sind als Verbraucher auf eine ausreichende Zufuhr an Energie durch bestimmte Energieträger angewiesen. Doch Energieträger sind kostbar, denn sie nützen den Verbrauchern nur dann etwas, wenn sie noch nicht „entwertet“ sind. Die naturgesetzliche Tendenz der Materie, Zustände höherer Entropie* anzunehmen, führt dazu, dass nutzbare Energie im Universum immer weiter abnimmt. Eine solche Entwertung eines Energieträgers ist beispielsweise das Verbrennen von Holz unter Freisetzung von CO2, während der umgekehrte Vorgang – das Wachsen von Pflanzen unter Aufnahme von CO2 – der Aufbau verwertbarer Energieträger ist (Fotosynthese). Der erste Vorgang ist unkompliziert, verläuft spontan und setzt Energie frei (exergoner* Vorgang), während der zweite Vorgang äußerst aufwändig reguliert ist und unter Energieaufnahme (Sonnenenergie) verläuft (endergoner* Vorgang).
Daher ist Sparsamkeit im Umgang mit nutzbarer Energie zwingend notwendig. Das zeigen auch aktuelle Bemühungen im Bereich der technischen Forschung und der Energiewirtschaft. So wird der Ausbau von Blockheizkraftwerken in Deutschland gefördert1, da diese Einrichtungen den Verlust der Wärmeenergie durch Abfluss in eine unerwünschte Richtung (Dissipation) minimieren sollen. Dazu dient das Prinzip der Kraft-Wärme-Kopplung, d. h. die Gewinnung von mechanischer Energie und Wärmeenergie in einem thermodynamischen* Vorgang. Ebenso sind moderne Fahrzeuge zunehmend mit Vorrichtungen versehen, die die Wiedergewinnung von Energie – z. B. beim Bremsen – ermöglichen sollen. Doch so sinnvoll diese Bemühungen auch sind, sie stoßen stets auf ein entscheidendes Problem: den hohen Aufwand für die Herstellung effizienter Vorrichtungen zur Einsparung von Energie. Denn bisher kann der Verlust von mindestens 50 % der wiedergewonnenen Energie durch Dissipation nicht verringert werden.2 Zudem ist die technische Realisierung solcher Vorrichtungen sehr anspruchsvoll. Es wundert daher nicht, dass der schwindelerregende Anschaffungspreis Verbraucher häufig davon abhält, zur Energieversorgung (Strom, Wärme) des eigenen Hauses ein Blockheizkraftwerk im Keller zu installieren.
Ausgehend von diesen leicht nachvollziehbaren Beobachtungen veranlasst die nähere Betrachtung biologischer Einrichtungen Wissenschaftler regelmäßig zum Staunen darüber, wie hier das „Energieproblem“ auf elegante und effiziente Art und Weise in verschiedenen Ausführungen gelöst wurde, um den Anforderungen des jeweiligen Organismus Rechnung zu tragen. Biologische Systeme zeichnen sich nicht nur durch einen erstaunlich sparsamen Umgang mit Rohstoffen aus, indem ein großer Teil der Abbauprodukte für den erneuten Aufbau biologischer Strukturen verwendet wird.
Sie weisen auch Vorrichtungen auf, die im Hinblick auf die Energieverwertung nahezu optimal sind.
|
Angesichts knapper Energieressourcen sind Organismen auf einen effizienten Energiestoffwechsel angewiesen. Während die Untersuchung der Atmungskette höherer Organismen schon vor über 40 Jahren zur Entdeckung von Proteinkomplexen mit Energiespareinrichtungen führte, wurden vergleichbare biochemische Funktionseinheiten bei anaeroben* Mikroorganismen erst vor gut einem Jahrzehnt entdeckt. Es handelt sich dabei um sogenannte Bifurkations-Enzyme, also biochemische Katalysatoren, die einen Elektronenfluss gabeln oder verzweigen. Diese können Elektronen gezielt auf zwei verschiedene Routen schicken – und das unter Kopplung energieverbrauchender mit energieliefernden Prozessen. Zuvor wurde angenommen, dass der Energiestoffwechsel der Anaerobier aufgrund des Fehlens solcher Enzyme ineffizient sei. Dies ist aus evolutionstheoretischer Sicht eine plausible Annahme, da anaerobe Organismen aufgrund ihrer vergleichsweise geringen Komplexität (z. B. kleines Genom) als gute Kandidaten für den „primitiven“ Ursprung des Lebens auf der Erde gelten. Zudem geht die Mehrheit der Forscher bisher davon aus, dass die ersten Organismen unter sauerstofffreien Bedingungen entstanden sind und daher (zunächst) anaerob lebten. Aufgrund von neueren Erkenntnissen stellt sich dieser Sachverhalt jedoch anders dar. Anaerobier weisen demnach ebenfalls hochkomplexe Bifurkations-Enzyme auf, die einen nahezu optimalen Energiestoffwechsel ermöglichen. Für evolutionstheoretische Erklärungen stellt dies ein schwerwiegendes Problem dar, da Leben schon auf der Stufe der geringsten Komplexität offenbar nahezu perfekt eingerichtet ist. Als Erklärungsversuche dienen unbegründete Behauptungen und hypothetische natürliche Vorgänge, für die aber bisher keine empirischen Belege vorliegen. Dagegen können die Bifurkations-Enzyme als klare Design-Indizien interpretiert werden. Es ist in diesem Zusammenhang nicht überraschend, dass einige Forscher in den eleganten chemischen Abläufen innerhalb der Bifurkations-Enzyme Vorbilder für innovative Techniken sehen.
|
|
|
|
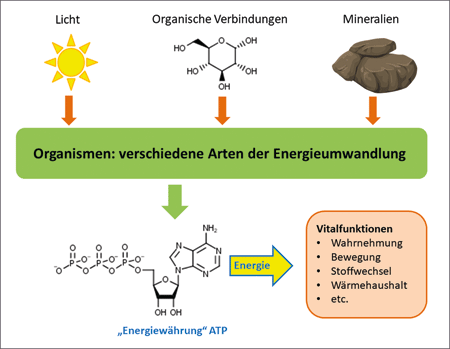
| Abb. 1: Schematische Darstellung des Prinzips des Energietransfers in der Biologie. Bis auf wenige Organismen speichern die Lebewesen die Energie von Primärträgern (Licht, organische Verbindungen, Mineralien) in der chemischen Verbindung ATP (Adenosintriphosphat). Diese „Energiewährung“ liefert die Energie für verschiedene Vitalfunktionen eines Organismus. |
„Energiekopplung“ bei aeroben Organismen
Ein grundlegendes Prinzip des Energiestoffwechsels aller Organismen beruht auf der Zwischenspeicherung der aufgenommenen Energie in energiereichen chemischen Verbindungen. Die meisten Organismen wandeln die aufgenommene Energie, sei es in Form von Licht, Mineralstoffen oder organischen Verbindungen, in die „Energiewährung“ Adenosintriphosphat (ATP) um (Abb. 1). Für diesen Vorgang werden überaus komplex organisierte Ensembles molekularer Konstruktionen, bestehend aus Multienzymkomplexen und einer Vielzahl an Kofaktoren, verwendet. Der Transfer der Energie erfolgt dabei durch physikalische Vorgänge (energetische Anregung, Ladungsfluss) im Zusammenspiel mit chemischen Reaktionen. Die in Form von ATP gespeicherte Energie wird später dazu genutzt, biochemische Reaktionen zu ermöglichen, die nicht spontan ablaufen.
Besondere Aufmerksamkeit verdienen hierbei molekulare Vorrichtungen, die offenbar so beschaffen sind, dass dissipative* Energieverluste beim Energietransfer minimiert werden. Bei Eukaryoten sind in diesem Zusammenhang viele Untersuchungen am Q-Zyklus, einem Teilvorgang der Atmungskette, durchgeführt worden. Der Q-Zyklus findet innerhalb des Komplexes III (Q-Cytochrom-c-Oxidoreduktase), einer mitochondrialen Protonenpumpe, statt.
|
|
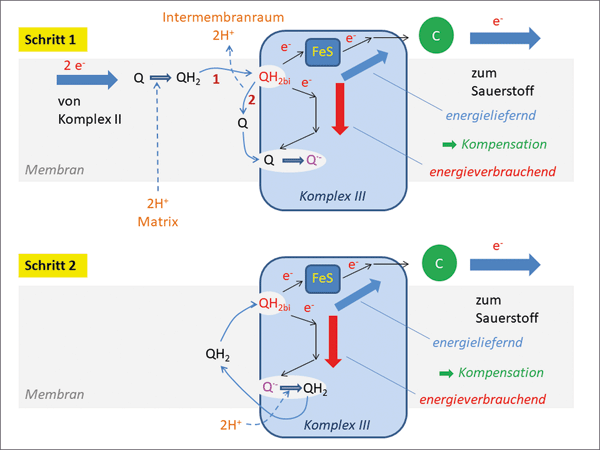
Abb. 2: Vereinfachtes Schema des Q-Zyklus in der mitochondrialen Atmungskette. Abkürzungen: Q Chinon, QH2 Ubichinon, QH2bi Ubichinon an der Stelle der Bifurkation, Q· - Semichinon, FeS Eisen-Schwefel-Protein, C Cytochrom. „Ecken“ im Elektronentransport stellen Zwischenstationen dar, die der Übersichtlichkeit wegen nicht dargestellt sind. Der Komplex III pumpt pro Übertragung von zwei Elektronen vier Protonen aus der mitochondrialen Matrix in den Intermembranraum. Die energieliefernde Reaktion ist die Reduktion des Cytochroms, sie kompensiert die energieverbrauchende Regeneration von QH2. |
Das Besondere an diesem Vorgang ist die getrennte Übertragung zweier Elektronen von demselben Molekül (Ubichinon) auf zwei verschiedene Elektronen-Akzeptoren* (ein anderes Chinon und ein Cytochrom c). Bildhaft gesprochen gabelt sich hier der Elektronenfluss, weswegen von einer „Elektronenbifurkation“ gesprochen wird (Abb. 2). Das Besondere an dieser molekularen Einrichtung ist die energetische Kopplung eines energieverbrauchenden (roter Pfeil) mit einem energieliefernden (blauer Pfeil) Vorgang. Auffällig ist dabei, dass die gekoppelten Energiebeträge annähernd gleich groß sind, sodass kaum Energie in Form von Wärme verloren geht (Details s. Kasten „Komplex III“).
|
Der Elektronentransfer innerhalb des mitochondrialen Komplexes III verläuft nach folgendem Prinzip (Abb. 2): Im ersten Schritt wird ein Elektron vom Ubichinon abgespalten und auf ein Chinon übertragen. Es resultiert dabei ein Radikalanion, auch als Semichinon bezeichnet, welches kurze Zeit „wartet“. Nach Durchlaufen einer zweiten entsprechenden Elektronenübertragung wird das Semichinon selbst zu einem Ubichinon reduziert und kann anschließend ebenfalls im gleichen Prozess oxidiert werden. Auffällig ist bei diesen Vorgängen die Energiebilanz: Während die Elektronenübertragung auf das Cytochrom c Energie freigesetzt, verbraucht die zweistufige Reduktion des Chinons zum Ubichinon Energie. Durch energetisches Koppeln der Vorgänge wird verhindert, dass die Energie, die bei der Reduktion des Cytochroms c freigesetzt wird, einfach verpufft. Stattdessen dient sie dazu, in einem energieverbrauchenden Vorgang die Hälfte der verbrauchten Ubichinone zu regenerieren. Dies stellt eine sehr effiziente Einrichtung bezüglich Material- und Energieverbrauch dar. Diese komplexe Konstruktion ist notwendig, um von einem Zwei-Elektronen-Transporter (Ubichinon) auf einen Ein-Elektron-Transporter (Cytochrom c) umzuschalten. Auf diese Weise wird eine effiziente Elektronenübertragung auf den Sauerstoff im benachbarten Proteinkomplex IV ermöglicht.
|
|
|
|
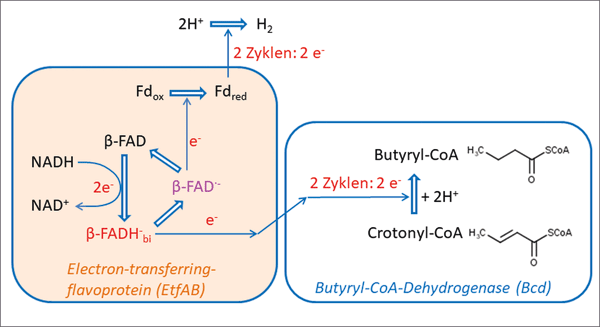
| Abb. 3: Vereinfachtes Schema der Abläufe im Bifurkations-Enzym EtfAB-Bcd. Abkürzungen: NADH Nicotinamidadenindinukleotid, b-FAD Flavin-Adenin-Dinucleotid, in der b-Untereinheit von EtfAB, b-FAD· - Semichinon, Fdox Ferredoxin-oxidierte Form, b-FADHbi reduziertes Flavin-Adenin-Dinucleotid an der Stelle der Bifurkation, Fdred Ferredoxin-reduzierte Form. „Ecken“ im Elektronentransport stellen Zwischenstationen dar, die der Übersichtlichkeit wegen nicht dargestellt sind. Der energieliefernde Prozess ist die Elektronenübertragung von NADH auf Crotonyl-CoA unter Bildung von Buturyl-CoA, sie kompensiert die energieverbrauchende Reduktion von Fdox. |
„Energiekopplung“ bei Anaerobiern
|
|
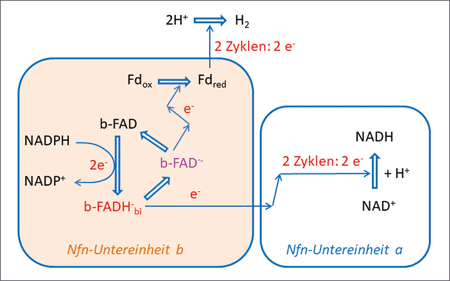
| Abb. 4: Vereinfachtes Schema der Abläufe im Bifurkations-Enzym NfnAB. Abkürzungen: NADH Nicotinamidadenindinukleotid, b-FAD Flavin-Adenin-Dinucleotid, in der Untereinheit b von NfnAB, b-FAD· - Semichinon, Fdox Ferredoxin-oxidierte Form, b-FADHbi reduziertes Flavin-Adenin-Dinucleotid an der Stelle der Bifurkation, Fdred Ferredoxin-reduzierte Form. „Ecken“ im Elektronentransport stellen Zwischenstationen dar, die der Übersichtlichkeit wegen nicht dargestellt sind. Der energieliefernde Prozess ist die Elektronenübertragung von NADPH auf NAD+ unter Bildung von NADH, sie kompensiert die energieverbrauchende Reduktion von Fdox. |
Während die Elektronenbifurkation im Komplex III von Eukaryoten schon 1975 von Mitchell3 beschrieben worden ist, wurde bis vor ungefähr zehn Jahren davon ausgegangen, dass anaerobe Organismen solche Vorrichtungen nicht aufweisen. Stattdessen wurde vermutet, dass Anaerobier einen ineffizienten Stoffwechsel aufweisen – eine Annahme, die aus evolutionstheoretischer Sicht4 naheliegend und sinnvoll erscheint, da Anaerobier als gute Kandidaten für den „primitiven“ Ausgangspunkt der evolutiven Höherentwicklung gelten. Zudem vertritt die Mehrheit der Forscher die Auffassung, dass die ersten Organismen in einer sauerstofffreien Umgebung entstanden sind, sodass sie zwangsläufig einen anaeroben Stoffwechsel betrieben haben müssten. In Anlehnung an diese Annahmen schreiben Buckel und Thauer (2018) im Rückblick auf die Ergebnisse früherer Untersuchungen und thermodynamischer Berechnungen zum Energiestoffwechsel von anaeroben Mikroorganismen:
„Aufgrund dieses Resultats und der Ergebnisse von anderen wurde [zuvor] angenommen, dass Anaerobier ineffiziente Energieverwerter sind […].“
Diese Sichtweise musste jedoch aufgrund von jüngeren Untersuchungen am Stoffwechsel von obligat* anaeroben Mikroorganismen revidiert werden. Denn es wurden bei mehreren Arten molekulare Einrichtungen nachgewiesen, die einen vergleichbar effizienten Energietransfer wie in aeroben Organismen durch Verwendung der Elektronenbifurkation ermöglichen. Die Entdeckung dieses Phänomens gelang Buckel und Thauer bei Untersuchungen an anaeroben Bakterien mit der Bezeichnung Clostridium kluveri. Diese eigenartigen Lebewesen können auf Trinkalkohol (Ethanol) und Essigsäure wachsen, chemischen Verbindungen also, die von vielen anderen Mikroorganismen als Abfallprodukte ausgeschieden werden und auf viele Organismen in höherer Konzentration toxisch wirken. C. kluveri wandelt die genannten Ausgangsverbindungen durch Fermentation* in Buttersäure und Capronsäure um und setzt dabei molekularen Wasserstoff frei (Seedorf et al 2008).
Lange Zeit war unklar, wie das Bakterium diese energetisch kostspielige Synthese bewerkstelligt, wobei vor allem die unter Atmosphärendruck stark endergone Reduktion von Wasserstoff energetisch besonders ins Gewicht fällt. Erst nachdem die Funktionsweise des aus drei Untereinheiten zusammengesetzten EtfAB-Bcd Enzymkomplexes (electron-transfering flavoprotein-butyryl-CoA dehydrogenase) beschrieben werden konnte, war das Rätsel gelöst: Durch elegante Kopplung energieverbrauchender mit energieliefernden Elektronenübertragungen innerhalb von EtfAB-Bcd ergibt sich eine thermodynamisch günstige Bilanz (∆G<0)* für den gesamten Vorgang (Herrmann et al. 2008).
Zudem weisen die gekoppelten Vorgänge ähnliche Energiebeträge auf und sind daher annähernd perfekt kompensatorisch, so dass es kaum zu einem Energieverlust kommt (Abb. 3; Details s. Kasten „EtfAB-Komplex“).
|
Im EtfAB-Komplex findet der Transfer von zwei Elektronen vom NADH auf Crotonyl-CoA statt (in zwei Zyklen). Der energieverbrauchende Vorgang ist hierbei die Reduktion von Ferredoxin (Eisen-Schwefel-Protein), das anschließend die Elektronen für den entstehenden Wasserstoff bereitstellt. Die hierfür erforderliche Energie wird aus der Reaktion von Crotonyl-CoA zu Butyryl-CoA und NADH (Nicotinamidadenindinukleotid) zu NAD+ gewonnen. Die Bifurkation geschieht hier an einem Flavin-Adenin-Dinukleotid (FAD), welches zunächst ein Elektron über eine Transportkette auf Crotonyl-CoA überträgt (exergone Reaktion) und anschließend ein Elektron auf ein Ferredoxin (endergone Reaktion). Da Crotonyl-CoA ein Zwei-Elektronen-Akzeptor ist, bedarf es zweier Zyklen innerhalb des Enzyms, bis ein Molekül Crotonyl-CoA vollständig zu Butyryl-CoA reduziert ist. Insgesamt sind an dieser Elektronenübertragungskaskade fünf Kofaktoren beteiligt. Die Reaktionsfolge ist in einem vereinfachten Schema wiedergegeben (Abb. 3).
|
Ein weiterer Typ von Enzymkomplex, dessen Funktionsweise die Elektronenbifurkation beinhaltet, ist die NADH-abhängige Ferredoxin(Fd) NADP+-Oxidoreduktase (Nfn), die aus verschiedenen Anaerobiern isoliert werden konnte. Besonders gründlich konnte ein Exemplar aus Thermotoga maritima anhand einer Röntgenkristallstrukturanalyse charakterisiert werden (Demmer et al. 2015). Bei T. maritima handelt es sich ebenfalls um ein obligat anaerobes Bakterium, das unter hyperthermophilen* Bedingungen lebt (ideale Temperatur 80 °C). Es gewinnt seine Energie aus der Umwandlung verschiedener organischer Verbindungen wie Zucker oder Zellulose in CO2 und H2. Auch im Fall der Nfn-Enzymkomplexe liegt eine annähernd perfekte Bilanz der Energieübertragung vor, obwohl sie einige bauliche Unterschiede zum EtfAB-Komplex aufweisen (Abb. 4; Details s. Kasten „Nfn-Komplex“).
|
Der aus T. maritima isolierte Nfn-Komplex besteht aus zwei Untereinheiten und katalysiert die Reduktion von Ferredoxin durch eine vermittelte Elektronenübertragung vom Kofaktor NADPH zu NADH. Der energieverbrauchende Vorgang ist hier ebenfalls die Elektronenübertragung auf das Ferredoxin, während der Elektronentransport zum NADH Energie liefert. Auch hier dient das reduzierte Ferredoxin schließlich der Synthese von H2. Die Bifurkation wird auch in diesem Enzymkomplex in Analogie zum EtfAB-Bcd-Komplex durch einen FAD-Kofaktor bewirkt. An der gesamten Elektronenübertragungskaskade sind acht Kofaktoren beteiligt. Die Elektronenübertragungen sind in einem vereinfachten Schema wiedergegeben (Abb. 4).
|
Interessanterweise gibt es auch Organismen, die die Elektronenbifurkation in umgekehrter Richtung nutzen, wie das Milchsäure-oxidierende Bakterium Acetobacterium wodii. A. wodii macht Gebrauch von einem EtfAB-Komplex, wobei hier die Elektronen vom Lactat und Ferredoxin ausgehend auf dem FAD an der Stelle der Gabelung zusammenfließen, um anschließend in einer Zwei-Elektronen-Übertragung zum NAD+ zu gelangen, sodass NADH gebildet wird (Weghoff et al. 2015). Ein solcher Vorgang wird konsequenterweise als Konfurkation bezeichnet. Die außerordentliche Effizienz von Enzymkomplexen, die von der Elektronenbifurkation Gebrauch machen, zeigt sich auch in der Nutzung anderer, überaus anspruchsvoller chemischer Prozesse für den Energiehaushalt. Acetogene Mikroorganismen wie Moorella thermoacetica verwirklichen die thermodynamisch äußerst ungünstige Reduktion von CO2 zu CO (Buckel & Thauer 2013) und manche Bakterien vermögen sogar den aromatischen Ring von Benzoaten oder Acetylen zu reduzieren – chemische Prozesse, die selbst im Labor nur durch Einsatz hochpotenter chemischer Reduktionsmittel realisiert werden können (Boll et al. 2016). Insgesamt sind bislang elf Bifurkations-Systeme experimentell nachgewiesen worden (Buckel & Thauer 2018).
|
|
Auffällige Konstruktionsmerkmale von Bifurkations-Enzymen
Bei näherer Betrachtung der Funktionsweise von Bifurkations-Enzymen stellt sich zunächst die Frage, wie die zwei Elektronen, die von demselben Molekül abgespalten wurden, in zwei energetisch entgegengesetzte Richtungen transportiert werden können – das eine Elektron in Richtung höheren elektrochemischen Potentials (energetisch bergab) und das andere in Richtung geringeren Potentials (energetisch bergauf). Denn gewöhnlich würde man erwarten, dass beide Elektronen energetisch bergab transportiert würden. Zudem erscheint es rätselhaft, dass es nach der ersten endergonen (energetisch bergauf) Elektronenübertragung nicht sofort zu einem Rücktransfer kommt. Eine wesentliche Voraussetzung hierfür ist eine äußerst präzise räumliche Anordnung der Kofaktoren innerhalb des Bifurkations-Systems. Zum einen sind die Kofaktoren so angeordnet, dass die jeweiligen Elektronen-Transportwege in entgegengesetzte Richtungen orientiert sind. Zum anderen sind die Abstände zwischen den Kofaktoren optimal für den Ladungstransport. Nach Zhang et al. (2017) würden selbst geringe Veränderungen der Distanzen zwischen den Kofaktoren den Elektronentransport innerhalb des von der Gruppe näher untersuchten Nfn-Komplexes signifikant beeinträchtigen:
|
|
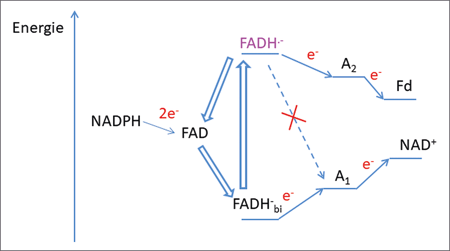
| Abb. 5: Schematische Darstellung der relativen Energien der an der Bifurkation beteiligten Komponenten (am Beispiel des Nfn-Komplexes). Zuerst werden zwei Elektronen vom NADPH auf FAD übertragen, sodass FADH-bi gebildet wird, welches dann ein Elektron auf den ersten Akzeptor (A1) überträgt (endergoner Vorgang). Das bei der Abspaltung gebildete Semichinon FADH- ist sehr energiereich und überträgt schnell ein weiteres Elektron auf den zweiten Akzeptor (A2). Der Übergang vom Semichinon auf A1 ist aufgrund der großen Energiedifferenz der beteiligten Orbitale kinetisch gehemmt. Der Fluss der Elektronen vom Semichinon zum Ferredoxin (Fd) verläuft energetisch bergab, währen der Fluss der Elektronen vom FADH-bi zum NAD+ energetisch bergauf verläuft. Die Energiebeträge der Übergänge sind ähnlich und kompensieren sich. |
„Dies bedeutet, dass eine Variation der Distanz des Elektronentransfers um wenige Angström (10-10 m) dazu führen würde, dass die Übertragungsrate [der Elektronen] sich um eine Größenordnung ändern würde.“
Weitere wichtige Größen sind passende physikochemische Parameter der Kofaktoren. Denn für einen reibungslosen Elektronentransport über mehrere Stationen müssen diese Parameter der Moleküle exakt aufeinander abgestimmt sein. Dieser Sachverhalt wird von Peters et al. (2018) wie folgt zum Ausdruck gebracht:
„Wie erreicht die Biologie die Ladungstrennung und Zustellung [von Elektronen]‚am richtigen Platz und zur richtigen Zeit‘? Die Antwort ist, dass die Natur sich die Physik der Elektronentunnel-Prozesse zunutze macht, welche mit [zunehmenden] Abständen zwischen Donoren und Akzeptoren exponentiell abfallen.“
Zusammenfassend kann also gesagt werden, dass in Bifurkations-Enzymen (Q-Zyklus, EtfAB, NfnAb) zugleich mehrere geometrische und physikochemische Parameter genau so eingestellt sind, dass eine optimale Elektronenübertragung im Verhältnis 1:1 und unter maximaler Energieausbeute auf zwei räumlich getrennte Kofaktoren realisiert wird. Die genau eingestellten physikalischen Parameter ermöglichen ferner eine extrem schnelle Übertragung des zweiten Elektrons vom Semichinon-Radikal (1011 s-1) auf den Akzeptor 2 (Abb. 5). Dies ist notwendig, da langlebige Radikale durch Nebenreaktionen zu Schäden am biologischen Material führen können (weitere Details s. Kastentext „Elektronentransfer in Bifurkations-Enzymen“).
Wissenschaftler, die sich eingehend mit Bifurkations-Systemen beschäftigt haben, werden oft dazu veranlasst, über die Architektur und die hochgradig effizienten Abläufe innerhalb dieser molekularen Entitäten zu staunen. So beschreiben Zhang et al. (2017) die Gesamtfunktion der Proteinkomplexe mit folgenden Worten:
 |
| Wissenschaftler staunen über die perfekte Architektur und effiziente Funktionsweise der Bifurkations-Enzyme. |
 |
|
„Obwohl die Elektronen-Bifurkation im Hinblick auf die Energieausbeute und die Zustellung der Elektronen nahezu perfekt funktioniert, handelt es sich um ein biologisches Energie-Transfer-Paradigma, das erst kürzlich breitere Aufmerksamkeit erfahren hat. […] Bemerkenswerterweise senden Bifurkations-Systeme ein Elektron [energetisch] bergauf und eines [energetisch] bergab mit ähnlichen Energiebeträgen, sodass der Gesamtprozess spontan verläuft, aber nicht verschwenderisch ist.“
Peters et al. (2018) verweisen auf die Exaktheit der Architektur und die Abstimmung der physikalischen Parameter von Kofaktoren als wichtige Voraussetzungen für optimale Abläufe in Bifurkations-Systemen:
„[Kofaktoren] verfügen über Architekturen, die eine Abspaltung des ersten Elektrons unter Ausbildung eines stark reduzierenden zweiten Donors ermöglichen, welche Schlüsselparameter (Abstand und Potenzialdifferenz) zur Kontrolle und Ausrichtung der zweiten Elektronenübertragung in die erforderliche Richtung verwenden.“
Bei der Betrachtung dieser biochemischen Systeme stellen sich ausgehend von der evolutionstheoretischen Denkweise folgende Fragen: Wie kann ein zukunftsblinder Prozess ein System hervorbringen, in dem annähernd perfekt kompensatorische Energietransfers ablaufen und einzelne Komponenten in optimaler Distanz für die Elektronenübertragung angeordnet sind? Wie kann ein ungerichteter Vorgang physikochemische Parameter derart adjustieren, dass Kontrolle und Ausrichtung möglich sind?
|
Die hier angesprochenen Elektronentunnelprozesse sind extrem schnelle Elektronenübertragungen, die auch thermodynamisch ungünstige Vorgänge ermöglichen können. Dazu bedarf es allerdings, wie bereits erwähnt, exakt eingestellter Abstände zwischen den beteiligten Molekülen. Zusätzlich müssen die am Elektronentransfer beteiligten Orbitale* der Moleküle energetisch gut zueinander passen. Bei zu großem energetischem Abstand zwischen den beteiligten Orbitalen kann die Übertragung deutlich verlangsamt werden (kinetische Hemmung).
Ein weiterer Umstand, der sicherstellt, dass es bei der Elektronenbifurkation nicht zu Rückreaktionen kommt, ist die besondere Beschaffenheit des Kofaktors, der an der Stelle der Bifurkation zum Einsatz kommt. Bei den bisher angeführten Kofaktoren Ubichinon (Komplex III, Atmungskette) und FAD ist die aufzuwendende Energie für die Abspaltung des ersten Elektrons deutlich größer als diejenige für die Abspaltung des zweiten Elektrons. Es verhält sich sogar so, dass nach der Abspaltung des ersten Elektrons (zur Übertragung auf den ersten Akzeptor) ein sehr energiereiches und kurzlebiges Radikal (Semichinon) entsteht, das sehr leicht das zweite Elektron abgibt (Abb. 5). Aufgrund der resultierenden großen Energiedifferenz zwischen dem Semichinon und dem ersten Akzeptor (A1) ist die Übertragung des zweiten Elektrons in die gleiche Richtung kinetisch gehemmt und es kommt zu einer schnellen Übertragung auf einen anderen Kofaktor (A2), dessen Orbitalenergie besser zu derjenigen des Semichinons passt. Auf diese Weise wird das zweite Elektron auf den Weg in die andere Richtung gelenkt.
|
|
|
Evolutive Entstehung von Bifurkations-Enzymen?
Angesichts des Vorkommens von offensichtlich hochkomplexen und effizient funktionierenden Enzymkomplexen bei Organismen, die landläufig als „primitiv“ bezeichnet werden und als gute Kandidaten für den Beginn der hypothetischen Höherentwicklung gelten, stellt sich die Frage nach deren Ursprung neu. Aktuell wird davon ausgegangen, dass die besten Kandidaten für den Beginn des Lebens Mikroorganismen sind, die kein ATP zur Zwischenspeicherung von Energie verwenden (Sousa et al. 2018), denn die Herstellung von ATP stellt eine zusätzliche Stufe der metabolischen Komplexität dar. Daher nehmen Sousa et al. an, dass der letzte universelle gemeinsame Vorfahre (last universal common ancestor, LUCA) ein Mikroorganismus war, der zu den acetogenen oder methanogenen Organismen gehörte, also zu Organismen, die die Kombination aus CO2 und H2 (neben einigen anderen chemischen Verbindungen) als Quelle für Energie und den für das zelluläre Baumaterial benötigten Kohlenstoff nutzen und Methan bzw. Essigsäure als Abfallprodukte ausscheiden. Solche Organismen machen Gebrauch vom Acetyl-CoA-Stoffwechsel (auch Wood-Ljungdahl-Weg), der auch ohne ATP betrieben werden kann. Allerdings erkennen Sousa et al. an, dass ein Stoffwechsel, der auf dem Elektronenfluss von H2 zu CO2 beruht und mit Bifurkations-Systemen ausgestattet ist, alles andere als leicht zu realisieren ist:
„Das Problem mit H2 [als Nahrungsquelle] im Kontext der frühen Evolution ist, dass Flavin-basierte Elektronen-Bifurkation ein elaborierter physiologischer Vorgang ist, der einer konzertierten Funktionsweise mehrerer komplexer Proteine im Sinne eines Energie-Stoffwechselwegs bedarf, um Ferredoxin reduzieren zu können (nötig für die Reduktion von CO2). Dies stellt ein typisches Henne-Ei-Problem dar; nämlich, wie CO2 durch H2 reduziert wurde, bevor es Proteine gab, die von der Bifurkation Gebrauch machten.“
Die Lösungsvorschläge für dieses Paradox, die in der besagten Veröffentlichung präsentiert werden, sind nicht überzeugend. Da ist zunächst die Rede von der „Serpentinisierung“* als möglicher Quelle für die Reduktion von CO2 zu Methan unter Beteiligung von Eisen(II)-Ionen als Elektronenquelle, wobei der genaue Mechanismus des Vorgangs „noch nicht bekannt“ sei. Da es sich bei der Serpentinisierung um einen rein geochemischen Prozess handelt, der weder fein abgestimmt noch exakt ausgerichtet ist, stellt sich allerdings die Frage, wie hier eine Brücke zu hochkomplexen Bifurkations-Systemen geschlagen werden kann. Als weiteres Argument wird die kürzlich in der Gruppe von Moran gemachte Beobachtung angeführt, dass CO2 an der Oberfläche von elementarem Eisen unter speziellen Bedingungen zu biologisch relevanten chemischen Verbindungen reduziert werden kann (Muchowska et al. 2017, Varma et al. 2018). Es wurde allerdings schon gezeigt, weshalb diese eleganten Labor-Experimente keinerlei Bezug zu einem plausiblen geochemischen Szenario haben (Schmidtgall 2019).
|
| Das „Wie“ als Frage nach einem konkreten Mechanismus eines evolutiven Vorgangs bleibt einmal mehr völlig unbeantwortet. |
|
|
Ein weiteres Problem ist das Vorkommen z. T. verschieden beschaffener, optimal justierter Bifurkations-Systeme in nahezu allen biologischen Spezies. Dieser Sachverhalt spricht klar gegen die gemeinsame Abstammung als mögliche Ursache für dieses Phänomen. Stattdessen wird einmal mehr konvergente* Evolution angenommen (Buckel & Thauer 2018):
„Zusammenfassend [kann gesagt werden, dass] die Bifurkations-Systeme ein schönes Beispiel für konvergente Evolution sind. Die drei Enzym-Komplexe, die keinerlei evolutionäre Beziehung aufweisen [Q-Zyklus, EtfAB, Nfn], haben offenbar ihre Reduktionspotentiale und Abstände [zwischen Kofaktoren] unabhängig voneinander adjustiert, um diesen Bifurkations-Prozess optimal durchführen zu können.“
Auch hier wird, wie so oft, ohne jegliche Begründung in den Raum gestellt, dass biochemische Einrichtungen irgendwie von selbst zu einer optimalen Funktionsweise gefunden haben sollen – und das auch noch mehrfach unabhängig mit vergleichbarem Ergebnis (Konvergenz). Auf Konkretisierungen und detaillierte Erklärungsversuche wird verzichtet, obwohl genau das erforderlich wäre. Das „Wie“ als Frage nach einem Mechanismus der Entstehung bleibt somit einmal mehr völlig unbeantwortet.
|
aerob: Als „aerob“ werden Organismen bezeichnet, die für ihren Stoffwechsel Sauerstoff benötigen. anaerob: Als „anaerob“ werden Organismen bezeichnet, die für ihren Stoffwechsel keinen Sauerstoff benötigen. Viele dieser Organismen sind empfindlich gegenüber Sauerstoff. Organismen, die keinen Sauerstoff vertragen, werden als „obligat anaerob“ bezeichnet. ∆G: Änderung der freien Enthalpie (-> freie Enthalpie) Dissipation: Verteilung der Energie in unerwünschte Richtungen (Streuung) in Form von Wärme. Elektronen-Akzeptor: Teilchen, das Elektronen aufnimmt. endergon: Ein Vorgang ist „endergon“, wenn Arbeit (thermodynamisch) in das betreffende System investiert werden muss. Die Arbeit kann dem System in Form von Wärme zugeführt werden oder indem die Ordnung des Systems erhöht wird. Solche Vorgänge sind durch einen positiven Betrag der Änderung der freien Enthalpie charakterisiert (∆G>0). Entropie: Maß für die Unordnung eines Systems (Zustandsgröße S). Wenn ein System aus einer bestimmten Anzahl an Teilchen besteht, können die Teilchen Zustände höherer Ordnung oder geringerer Ordnung annehmen. Wenn der Zustand der Teilchen von einem geordneten in einen weniger geordneten wechselt, steigt die Entropie des Systems, d. h. ∆S>0 (in J/K · mol). exergon: Ein Vorgang ist „exergon“, wenn ein System dabei (thermodynamisch) Arbeit leistet. Solche Vorgänge sind durch einen negativen Betrag der Änderung der freien Enthalpie charakterisiert (∆G<0). Fermentation: Umwandlung von organischen Verbindungen in Gase, Säuren oder Alkohol durch Mikroorganismen. freie Enthalpie (G): Größe, die den Energiegehalt (Zustand) eines Systems beschreibt. Das Maß für die Arbeit, die ein System leistet oder aufnimmt, ist die Änderung der freien Enthalpie ∆G (in kJ/mol). Ein negativer Betrag von ∆G bedeutet, dass das System Arbeit leistet. Für alle Naturvorgänge, die spontan verlaufen, gilt ∆G<0. Wird dem System Energie zugeführt, ist ∆G>0. Für Naturvorgänge, die unter Investition von Energie verlaufen, ist ∆G positiv. hyperthermophil: Als „hyperthermophil“ gelten Organismen, die bevorzugt unter besonders hohen Temperaturen leben. konvergente Evolution: Als „konvergent“ werden evolutive Abläufe bezeichnet, die unabhängig von Verwandtschaftsverhältnissen zu sehr ähnlichen biologischen Einrichtungen führen sollen. So gilt (im Rahmen der Synthetischen Theorie) das Echoortungssystem bei Fledermäusen und Delphinen als ein Beispiel für konvergente Evolution, da beide Lebewesen aufgrund ihrer großen morphologischen Unterschiede für stammesgeschichtlich sehr weit voneinander entfernt gehalten werden, sodass gemeinsame Abstammung nicht die Ursache des ähnlichen Merkmals sein kann. obligat: ausschließlich. Orbital: Begriff für einen Raum in einem Atom oder Molekül, in dem Elektronen sich mit großer Wahrscheinlichkeit aufhalten. Serpentinisierung: Umwandlung des eisen- und magnesiumhaltigen Minerals Brucit (durch Reaktion mit SiO2) zu Serpentin unter Bildung von Wasserstoff und Magnetit (Fe3O4) in ca. 400 °C heißen Quellen („Schwarzen Rauchern“) am Meeresboden. Es wird angenommen, dass die Bildung des Wasserstoffs die Folge der Umwandlung des Brucits ist. Thermodynamik: Auch als „Wärmelehre“ bezeichnet, d. h. die Lehre über das Verhalten von Vielteilchensystemen im Hinblick auf die Zuführung oder Abführung von Energie (Wärmeenergie, mechanische Energie). Zentrale Bedeutung haben Zustandsgrößen, die den Energiegehalt des Systems beschreiben, und Änderungen von Zustandsgrößen, die beim Energietransfer auftreten.
|
|
|
Bifurkations-Enzyme als Vorbild für technische Innovationen
Nicht selten werden in der Fachliteratur zur Beschreibung der Bifurkations-Enzyme Attribute wie „genial“ oder „elegant“ gebraucht. Das ist nicht weiter verwunderlich, denn die Chemie, die im Stoffwechsel der betreffenden Organismen verwirklicht wird, lässt jeden Chemiker staunen. Daher sehen einige darin eine Quelle für zukunftsweisende, bioinspirierte chemische Reaktionsfolgen, die möglicherweise auch für technische Anwendungen genutzt werden könnten:
„Insgesamt erklärt der hier etablierte theoretische Rahmen […] die Logik der Elektronenbifurkation, die die Energieumwandlung und -einsparung im Nfn-Komplex ermöglicht. Solche Vorgänge verweisen auf bioinspirierte Schemata zur Durchführung von Multielektronen-Redoxchemie und etablieren die Route für die Suche nach weiteren Bifurkations-Netzwerken in der Natur“ (Zhang et al. 2017).
„Ein fundamentales Verständnis der unfehlbaren Genauigkeit der Mechanismen und der von Elektronen-Bifurkations-Enzymen aufgeprägten eleganten Kontrolle verweist auf wichtige Prinzipien für das Design von Redox-Katalysatoren zur Energieumwandlung“ (Peters et al. 2018).
Offenbar zeigt sich auch hier, dass die Anerkennung des unübersehbaren Designs in der Natur ein fruchtbarer Ansatz für anwendungsorientierte Forschung ist. Im Gegensatz dazu birgt der Evolutionsgedanke keinerlei Potenzial für innovative Techniken.
|
|
Fazit
Insgesamt wird deutlich, dass die hier diskutierten Bifurkations-Enzyme hochgradig komplexe molekulare Einrichtungen darstellen, die über eine exakte Architektur und fein abgestimmte physikochemische Parameter der Komponenten verfügen. Diese Voraussetzungen ermöglichen eine praktisch perfekte Effizienz im Hinblick auf den Energietransfer bzw. die Energieersparnis. Es erscheint mehr als naheliegend, dass solche Vorrichtungen in Analogie zu Energiespareinrichtungen in Heizkraftwerken oder Fahrzeugen als Meisterwerke der Ingenieurskunst angesehen werden können. Ihre Entstehung durch einen ungerichteten Naturprozess (Zufall und Notwendigkeit) erscheint dagegen abwegig.
Das Vorkommen solcher molekularer Konstruktionen z. T. verschiedener Ausprägung in nahezu allen Typen von Lebewesen stellt nicht nur ein schwerwiegendes Problem für die Frage nach der Entstehung erster Lebensformen dar, sondern spricht auch gegen die gemeinsame Abstammung als mögliche Ursache für das Vorhandensein dieser Proteine. Die Annahme, es handle sich hierbei um ein Beispiel für konvergente Evolution, stellt eine ad-hoc-Behauptung dar, für die es keine belastbaren empirischen Belege gibt.
|
Anmerkungen
1 Kraft-Wärme-Kopplungs-Gesetz seit 2002, Erneuerbare Energien-Gesetz seit 2009, Steuervergünstigungen seit 2012
2 https://www.energie-lexikon.info/rekuperation.html, abgerufen am 13. 2. 19
3 Peter D. Mitchell wurde dafür 1978 mit dem Chemie-Nobelpreis ausgezeichnet.
4 Mit „evolutionstheoretisch“ ist die klassische Synthetische Evolutionstheorie gemeint.
Alle Hervorhebungen in den Zitaten stammen vom Autor dieses Artikels.
|
Literatur
- Boll et al. (2016)
- Structure and function of the unusual tungsten enzymes acetylene hydratase and class II benzoyl-coenzyme A reductase. J. Mol. Microbiol. Biotechnol. 26, 119–127.
- Buckel W & Thauer RK (2013)
- Energy conservation via electron bifurcating ferredoxin reduction and proton/Na+ translocating ferredoxin oxidation. Biochimica et Biophysica Acta 1827, 94–113.
- Buckel W & Thauer RK (2018)
- Flavin-based electron bifurcation, a new mechanism of biological energy coupling. Chem. Rev. 118, 3862−3886.
- Demmer et al. (2015)
- Insights into Flavin-based electron bifurcation via the NADH-dependent reduced ferredoxin: NADP oxidoreductase structure. J. Biol. Chem. 290, 21985–21995.
- Herrmann et al. (2008)
- Energy conservation via electron-transferring flavoprotein in anaerobic bacteria. J. Bacteriol 190, 784–791.
- Mitchell P (1975)
- The protonmotive Q cycle: a general formulation. FEBS Letters 59, 137–139.
- Muchowska KB et al. (2017)
- Metals promote sequences of the reverse Krebs cycle. Nat. Ecol. Evol. 1, 1716–1721.
- Peters JW et al.(2018)
- On the nature of organic and inorganic canters that bifurcate electrons, coupling exergonic and endergonic oxidation – reduction reactions, Chem. Comm. 54, 4091–4099.
- Seedorf H et al. (2008)
- The genome of Clostridium kluveri, a strict anaerobe with unique metabolic features. Proc. Natl. Acad. Sci.105, 2128–2133.
- Schmidtgall B (2019)
- Schritte zum Leben ohne Enzyme? https://www.genesisnet.info/schoepfung_evolution/n264.php.
- Sousa FL et al. (2018)
- Native metals, electron bifurcation, and CO2 reduction In early biochemical evolution, Curr. Opin. Microbiol. 43, 77–83.
- Varma SJ et al. (2018)
- Native iron reduces CO2 to intermediates and endproducts of the acetyl-CoA pathway. Nat. Ecol. Evol. 2, 1019–1024.
- Weghoff MC et al. (2015)
- A novel mode of lactate metabolism in strictly anaerobic bacteria. Environ. Microbiol. 17, 670–677.
- Zhang P (2017)
- Electron Bifurcation: Thermodynamics and Kinetics of Two-Electron Brokering in Biological Redox Chemistry. Acc. Chem. Res. 50, 2410–2417.
|
|  |