 |
Muschelaugen – Hightech im Kleinen
Zwei Netzhäute und ein Spiegelteleskop aus quadratischen Minikristallen in einem Millimeter-Auge
von Henrik Ullrich
Studium Integrale Journal
25. Jahrgang / Heft 2 - Oktober 2018
Seite 105 - 107
|
 |
Zusammenfassung: Wie sehen Muscheln im trüben Wasser? Lange schon war bekannt, dass Kammmuscheln über spezielle Reflexionsspiegel in ihren nur 1 mm großen Linsenaugen verfügen, um die Lichtausbeute zu optimieren. Neue Mikroskopiertechniken legen weitere geniale Details des Aufbaus und der Funktion dieser Augen offen, die unerwartet große Ähnlichkeiten mit Augen anderer Tierklassen aufweisen.
|
|
 |
| Abb. 1: A Kammmuschel (Pecten maximus) mit zahlreichen Augen am Mantelrand; der Pfeil zeigt auf eines davon. B Fluoreszenzmikroskopie des Linsenauges der Kammmuschel. (i) Hornhaut, (ii) Linse, (iii) vordere Netzhaut, (iv) hintere Netzhaut, (v) Reflektorspiegel; gelber Pfeil: gerader Lichteinfall. (Aus Pabner et al. 2017, mit freundlicher Genehmigung) |
|
Einleitung
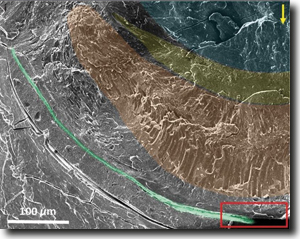 |
| Abb. 2: Kryo-Elektronen-Mikroskopie der Netzhaut und des Spiegels des Pecten-Auges. Linse (blau), vordere Netzhaut (gelb), hintere Netzhaut (braun), Reflektorspiegel (grün). Gelber Pfeil: gerader Lichteinfall. (Aus Pabner et al. 2017, mit freundlicher Genehmigung) |
|
Organismen verfügen in Abhängigkeit von ihrem Lebensraum und ihrer Lebensweise über eine faszinierende Vielfalt von optischen Apparaten und Einrichtungen, um das Licht und die damit übertragenen Informationen aus der Umgebung ihres Lebensraums wahrnehmen und verarbeiten zu können. Zentrale Elemente in der Kette biologischer Bausteine stellen Linsen und bei wenigen Tieren auch hinter der Netzhaut gelegene Spiegel dar, mit denen das ankommende Licht gebündelt und punktgenau auf die Netzhaut (Retina) mit ihren Lichtsinneszellen (Rezeptoren) reflektiert und fokussiert wird.
Das der biologischen Forschung bereits bekannte eindrucksvolle Repertoire entsprechender optotechnischer Lösungen ist nun um ein weiteres Highlight erweitert worden. Gefunden wurde es bei der Großen Pilgermuschel (Pecten maximus) aus der Ordnung der Kammmuscheln, zu deren bekanntesten Arten auch die Jakobsmuscheln gehören. Am Rand ihrer Gehäuse finden sich bis zu 200 Augen, die eine Größe von jeweils 1 mm aufweisen. Damit ist ein Sichtumfeld von ca. 250 ° abgedeckt. Seit den 1960er-Jahren wusste man bereits, dass in diesen Augen ein biologischer Hohlspiegel genutzt wird, der hinter der Netzhaut gelegen ist, um das in das Auge eintretende und die Netzhaut zunächst durchscheinende Licht auf die Netzhaut zurückzuwerfen und zu fokussieren. Die meisten Organismen, z. B. Wirbeltiere oder Tintenfische, nutzen für die Bündelung der Lichtstrahlen dagegen Linsen, die vor der Netzhaut gelegen sind. Einige Krustentiere und wenige Tiefseefische nutzen wie die Kammmuscheln zusätzlich zu ihren Linsen noch solche Spiegel.
 |
| Der Spiegel besteht aus Kristallen von Guanin, das in der Zelle an anderer Stelle als Baustein der DNA verwendet wird – eine ausgefallene Mehrfachnutzung eines Moleküls. |
 |
|
Ein Einzelauge der Kammmuschel besteht aus einer schwach lichtbrechenden Linse, dem Hohlspiegel und einer höchst ungewöhnlich aufgebauten, zweischichtigen Netzhaut. Die molekulare Feinstruktur des Spiegels und damit auch dessen Funktion im Zusammenspiel mit der doppelschichtigen Netzhaut bei diesen Muscheln konnte durch die Arbeitsgruppe von Pabner et al. (2017) unter Nutzung der CEM (cryo-electron microscopy) – einer Variante der Elektronenmikroskopie – aufgedeckt werden. Ihre Ergebnisse sorgten in mehrfacher Hinsicht für großes Erstaunen.
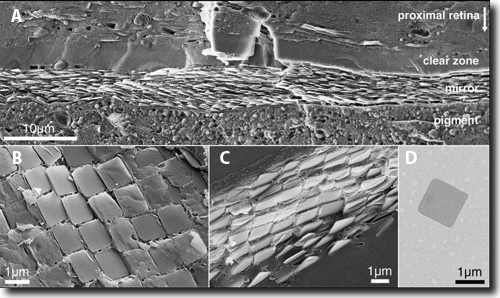 |
Abb. 3: Hochaufgelöste Kryo-Elektronen-Mikroskopie des Spiegels im Auge der Muschel. Beachte Maßstabsbalken! A 20-30 übereinandergelagerte Schichten von Guaninkristallen, B, C fliesenartige Anordnung der Guaninkristalle in einer Schicht, D einzelnes quadratisches Guaninkristall. (Aus Pabner et al. 2017, mit freundlicher Genehmigung) |
|
Der Spiegel – wir reden hier von einem Auge mit der Größe von 1 Millimeter – besteht aus 20-30 übereinander gelagerten Einzelschichten, deren biochemische Substanz aus Guaninkristallen gebildet wird. Dieses Molekül wird in der Zelle an anderer Stelle als Baustein der DNA, des molekularen Trägers der genetischen Information, verwendet. Die einzelnen flachen Kristalle sind quadratisch und lagern sich flächig aneinander und übereinander. Es ist ungeklärt, welche intrazellulären und biochemischen Mechanismen dafür verantwortlich sind, dass Guanin erstens diese flächigen exakten Quadratkristalle mit einer Kantenlänge von ca. 1,23 µm und einer Dicke von 74 ± 9 nm ausbildet, und zweitens, wie sie sich – wie Fliesen in einem Bad – flächig anordnen und bis zu 30 mal übereinander dicht gepackt werden (gesamte Dicke ca. 2,5 µm). Ein einzelnes Guaninkristall ist durchsichtig. Übereinander geschichtet reflektieren diese Kristallverbände jedoch ausgewählte Wellenlängen des ankommenden Lichts (nach den Prinzipien der Interferenz) mit einer Wellenlänge von ca. 500 nm im Bereich der Farben grün und blau, was dem Hauptanteil des ankommenden Lichts im Lebensraum der Muscheln entspricht. Besonders zu erwähnen ist noch, dass diskrete Abweichungen von einer exakten sphärischen Ausformung des Hohlspiegels dazu führen, dass die reflektierten Lichtanteile nicht auf einen, sondern zwei Brennpunkte zurückgeworfen werden.
Die Autoren fassen diesen Teil der Ergebnisse wie folgt zusammen: „Die hierarchische Organisation des Spiegels der Pilgermuschel ist fein abgestimmt für die Bildgebung, angefangen von den Guaninkristallen, den Komponenten im Nanobereich, bis zur gesamten Form der Spiegels im Millimeterbereich.“
|
|
Ungewöhnlich: zwei Netzhautschichten
|
| Der Mythos, Augen seien evolutionäre Fehlkonstruktionen, kann durch ungezählte Untersuchungsergebnisse als widerlegt gelten. |
|
|
Die Netzhaut des einzelnen Muschelauges verfügt, wie bereits angesprochen, in ungewöhnlicher Weise über zwei Schichten von Sinneszellen, die jeweils in exakter Distanz zum Spiegel entsprechend den beiden Brennweiten angeordnet sind. Unter Nutzung einer röntgenbasierten Mikro-Computertomographie des gesamten Auges mit zusätzlicher 3D-Modellierung und mathematischer Analyse der möglichen Lichtachsen wurde gezeigt, dass die komplexe 3D-Morphologie des gesamten Auges dazu beiträgt, dass Bilder auf beiden Netzhautschichten entstehen. Dabei zeichnet sich auch eine funktionelle Differenzierung der Netzhautanteile in Abhängigkeit vom Eintrittswinkel, der Helligkeit des Lichtes und der Position des Lichtobjektes in Bezug zur Muschel ab.
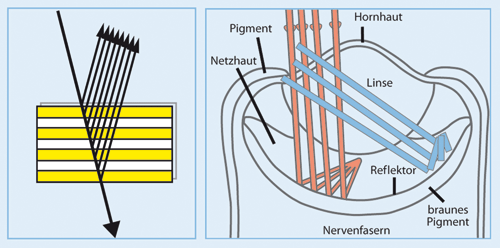 |
| Abb. 4: A Prinzip der Interferenz: Die auf eine Multilayer-Schicht eines Reflektors eintreffenden Lichtwellen mit unterschiedlichen Wellenlängen werden an den einzelnen Schichten reflektiert und übereinander gelagert. Dabei werden in Abhängigkeit vom Abstand der einzelnen reflektierenden Schichten bestimmte Wellenlängen verstärkt, andere werden ausgelöscht. B Schema der Reflexion und der Interferenz von Lichtwellen in Abhängigkeit vom eintreffenden Winkel der Lichtwellen auf die Multilayer-Schicht des Reflektorspiegels der Kammmuschel. Es konnte eine funktionelle Differenzierung der vorderen und hinteren Netzhaut sowie der jeweiligen zentralen und peripheren Anteile der Netzhaut in Abhängigkeit vom Eintrittswinkel, der Helligkeit des Lichtes und der Position des Lichtobjektes in Bezug zur Muschel nachgewiesen werden. Weitere Details siehe Text. (Nach Wikimedia commons, geändert) |
|
Die proximale (vordere) Netzhaut bildet besser Lichtobjekte unterhalb, die distale (hintere) Netzhaut besser Lichtobjekte oberhalb der Muschel ab. Das schärfste Sehen gelingt mittels der proximalen Netzhaut in ihren äußeren Anteilen und mittels der distalen Netzhaut in ihren zentralen Anteilen. Die Sinneszellen der letzteren reagieren auch deutlich schneller auf dunkle und sich bewegende Objekte. Die proximale Retina hingegen reagiert langsamer, aber dafür sensitiver auf sehr lichtschwache Objekte.
Diese funktionelle Aufteilung der Netzhautregionen erinnert an ähnliche Befunde der funktionsspezifischen Differenzierung der Netzhaut bei vielen anderen Organismen (z. B. spezialisierte Reihen von Ommatiden bei Insektenaugen zur Wahrnehmung von polarisiertem Licht oder die Verteilung von Stäbchen und Zapfen auf der Netzhaut der Säugetiere für das Hell-Dunkelsehen bzw. das Farbensehen).
Die Autoren fassen zusammen: „Auf der höchst möglichen hierarchischen Ebene erscheint der komplexe 3D-Aufbau des Augenspiegels der Muschel so abgestimmt zu sein, dass das Licht aus einem weiten Sichtfeld auf zwei Netzhäute in unterschiedlichem Abstand zu deren Oberflächen fokussiert werden kann. … Die Kristallmorphologie, die Mehrschichtstruktur und der 3D-Aufbau des Augenspiegels der Muschel sind fein abgestimmt, um funktionelle Bilder auf ihren beiden Netzhäuten zu erzeugen.“
|
|
Diskussion
Der Mythos, dass Augen evolutionäre Fehlkonstruktionen seien und die Narben ihrer evolutionären Geschichte trügen, kann durch die ungezählten Untersuchungsergebnisse zur Physiologie und zum Aufbau der Augen zahlreicher Organismen, einschließlich des Menschen, als widerlegt gelten. Eine evolutionäre Erklärung der Entstehung von Augen und der in ihnen verwirklichten optotechnisch, biochemisch und anatomisch raffinierten Baupläne ist bislang nicht einmal in Ansätzen möglich. Dazu kommt, dass es ein enormes Ausmaß an morphologischen und funktionellen Konvergenzen gibt – teils nahezu identische Lösungen für den Bau und die Funktion von Augen nicht verwandter Arten (z. B. Jonasova & Kozmik 2008). Auch dieser Befund spricht deutlich für einen intelligenten und damit schöpferischen Ursprung dieser Organe als vernünftige Alternative zur ziellosen und flickwerkartigen Evolution.
|
Literatur
- Jonasova K & Kozmik Z (2008)
- Eye evolution: Lens and cornea as an upgrade of animal visual system. Seminars in Cell & Dev. Biol. 19, 71-81.
- Pabner BA, Taylor GJ, Brumfeld V, Gur D & Shemes M (2017)
- The image-forming mirror in the eye of the scallop. Science 358, 1172-1175.
|
|
|  |
