|
Ribonukleinsäure (RNA), Proteine und Fettsäuren sind grundlegende Makromoleküle aller lebenden Zellen. Die Frage nach ihrem Ursprung ist daher eine der zentralen Fragen der chemischen Evolution, welche der Entstehung des Lebens vorausgegangen und diese ermöglicht haben soll. Ein wesentliches Problem ist dabei, dass nach aller chemischen Erfahrung sehr unterschiedliche chemische Bedingungen für die Entstehung der verschiedenen Biomoleküle benötigt werden. Kürzlich hat die Forschungsgruppe um John Sutherland (Patel et al. 2015) ein neues Modell als potentielle Lösung für dieses Problem vorgestellt. Sutherland beschreibt darin ein hypothetisches Szenario für die Entstehung von Vorläufern aller drei Molekülklassen – RNA, Proteine und Fettsäuren – in einem einzigen Ansatz. Konnte man dem Rätsel der Lebensentstehung durch diese Arbeit einen entscheidenden Schritt näher kommen?
 |
Für den Photoredoxzyklus wird
UV-Licht benötigt. Das eingesetzte
UV-Licht ist allerdings schädlich für
das gewünschte Endprodukt. |
 |
|
Sutherland fasst in dieser Arbeit Erkenntnisse aus früheren Untersuchungen zusammen und fügt neuere Erkenntnisse und Überlegungen an. Über eine der Pionierarbeiten (Powner et al. 2009) berichtete bereits Binder (2009) in Studium Integrale Journal. In der Arbeit von Powner et al. wurde ein möglicher präbiotischer Entstehungsweg für aktivierte Pyrimidinribonukleotide beschrieben – einem Baustein der Nukleinsäuren. In einer weiteren Arbeit konnte Sutherland zeigen (Ritson & Sutherland 2012), dass durch kupferkatalysierte Photoredoxchemie über Homologisierungsreaktionen aus der Vorläufersubstanz Cyanwasserstoff (HCN) C2- und C3-Zucker entstehen können, die als Ausgangsstoffe für die 2009 beschriebene Chemie gebraucht werden. Die von Ritson & Sutherland beschriebenen chemischen Reaktionen werfen im Kontext einer Chemie der Lebensentstehung allerdings viele Fragen auf:
- Die grundlegendste Anfrage ist, unter welchen Bedingungen der beschriebene Photoredoxzyklus überhaupt ablaufen kann. Die vorgestellte Photoredoxchemie funktioniert nämlich nur, wenn die beteiligten Komponenten räumlich isoliert und konzentriert sind, dagegen nicht in starker Verdünnung oder in Gegenwart anderer Chemikalien. Die beiden letzteren ungünstigen Randbedingungen müssen aber für eine Ursuppe angenommen werden.
- Für den Photoredoxzyklus wird UV-Licht (254 nm) benötigt. Das eingesetzte UV-Licht ist allerdings schädlich für das gewünschte Endprodukt. Es führt unter anderem zur Bildung von Pyrimidindimeren und damit zur massiven Schädigung von RNA und DNA. Die Nukleinsäuren können dann keine dynamischen dreidimensionalen Strukturen ausbilden und auch keine katalytischen Funktionen übernehmen oder genetische Informationen speichern. Diese Bedingungen wären also in jedem Fall inkompatibel mit einer RNA-Welt. Zudem ist die Fragestellung der Entstehung von RNA in den beschriebenen Arbeiten noch nicht einmal tangiert, da darin nur einige Vorläufer möglicher Ausgangsstoffe betrachtet werden. Die Polymerisation dieser Bausteine ist ein eigenes, zusätzliches und aus rein naturalistischer Perspektive vollkommen ungelöstes Rätsel (Schmidtgall 2013).
-
Die Frage nach der Plausibilität des Vorhandenseins großer Mengen von Cyanwasserstoff und weiterer benötigter Bausteine wie Phosphate muss geprüft werden.
In weiteren Arbeiten hat Sutherland gezeigt, dass zudem Schwefelwasserstoff eingeführt werden muss, um Vorläufer von Aminosäuren herzustellen (Ritson & Sutherland 2013). Durch diese Vorgehensweise konnten Zwischenprodukte für die Synthese von drei proteinogenen Aminosäuren synthetisiert werden. Hier soll nur auf die Synthese eines Zwischenprodukts für die Synthese der Aminosäure Valin eingegangen werden. Wie aus Abb. 1 ersichtlich, muss das Molekül 11 in mehreren, aufeinanderfolgenden Reaktionen unter jeweils unterschiedlichen Bedingungen synthetisiert werden. Für die Synthesen der Zwischenprodukte der beschriebenen Aminosäuren werden typischerweise unterschiedliche Randbedingungen für die chemische Reaktionen benötigt. Die Syntheseschritte und die dazu notwendigen Randbedingungen von 11 werden im Folgenden betrachtet; sie sind ein Kernbestandteil der neuesten Arbeit von Sutherland (Patel et al. 2015).
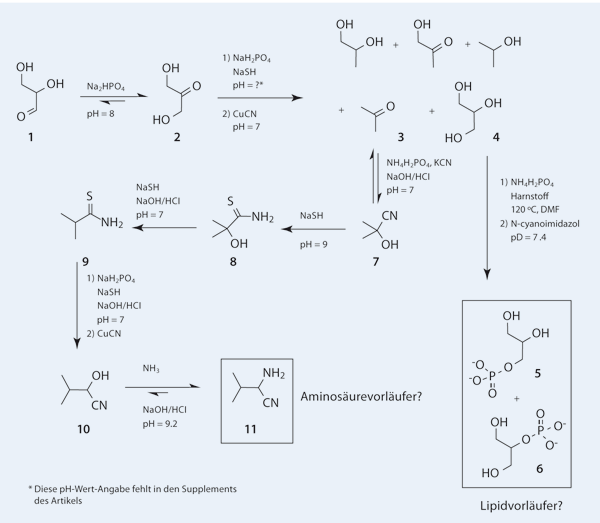 |
| Abb. 1: Ausschnitt aus dem in Patel et al. (2015) beschriebenen Netzwerk. |
|
Wenige neue Erkenntnisse
Im Zentrum der aktuellen Arbeit (Patel et al. 2015) steht die Umsetzung von Glycerinaldehyd (1 in Abb. 1) zu Dihydroxyaceton (2). Diese ist bei leicht basischem pH-Wert thermodynamisch begünstigt. Ausgehend von 2 stellen die Autoren ein Reaktionsnetzwerk vor, bei dem verschiedene Aminosäurevorläufer und Lipidvorläufer synthetisiert werden. Bei den vorgeschlagenen Lipidvorläufern handelt es sich um Glycerolphosphate (5 und 6). In Lipiden, die am Aufbau von Zellmembranen beteiligt sind, sind ähnliche Moleküle mit Fettsäuren verestert. Für die Funktion von Lipiden in Zellmembranen ist ihr ampiphiler Charakter entscheidend, d. h. sie „lieben“ sowohl Wasser als auch wasserabweisende Öl-artige Medien. Fettsäuren könnten diese Funktion prinzipiell erfüllen, aber Glycerolphosphate nicht. Glycerolphosphate erscheinen in diesem Licht eher überflüssig und bringen keinen Fortschritt für die Erklärung der Herkunft einfacher amphihiler Moleküle wie Alkylphosphate, Alkylsulfate oder Fettsäuren (Ruiz-Mirazo et al. 2014). Die Einschätzung von Glycerolphosphaten als Lipidvorläufer ist zwar sachlich nicht falsch, ein entscheidender Teil fehlt jedoch (die lipophile Kette) und derzeit gibt es auch keine Ansatzpunkte in Sutherlands Chemie, die eine Entstehung langer Alkylketten plausibel machen könnten. Für Moleküle mit amphiphilen Eigenschaften sind diese Ketten aber unbedingt notwendig.
Im gesamten Reaktionsnetzwerk, das Sutherland et al. konzipiert haben, ist die Gegenwart von löslichem Phosphat unabdingbar. Dieses erfüllt mehrere grundlegende Funktionen: Es verhindert starke Schwankungen des pH-Werts, katalysiert gewünschte Reaktionen und stabilisiert Verbindungen, die sonst zerfallen würden. Eine signifikante Phosphatquelle auf der Erde ist Apatit, ein Calciumphosphat-Mineral, das aber unreaktiv und schlecht wasserlöslich ist. Daher wirft eine mögliche Herkunft von löslichen Phosphaten schwerwiegende Fragen auf, und eine plausible Phosphatquelle ist nicht bekannt (Ruiz-Mirazo et al. 2014).
Neben vielen Fragen bezüglich der Anwesenheit bestimmter chemischer Verbindungen muss aber auch das Gesamtkonzept von Sutherland grundsätzlich hinterfragt werden.
Ein fragwürdiger Grundansatz
Für Szenarien der präbiotischen Chemie, also chemischer Synthesen, die zur Erzeugung von Stoffen zum Aufbau erster lebender Systeme genutzt werden können, ist die Konzeption möglichst einfacher und vor allem einheitlicher Reaktionsbedingungen erforderlich („Eintopfreaktionen“). Komplizierte Mechanismen, räumliche Trennungen und passende Abfolgen sind ohne Planung und Vorüberlegung wenig plausibel. Daher sind für die präbiotische Chemie Eintopfreaktionen viel wahrscheinlicher und einfacher zu begründen als komplexe, räumlich abgetrennte Reaktionen, deren Produkte zunächst in Isolation angereichert werden müssen und dann unter anderen Reaktionsbedingungen mit Stoffen weiter reagieren, die wiederum unter anderen speziellen Bedingungen erzeugt worden sind. Ein klassisches Beispiel für einen präbiotischen Eintopfversuch ist das bekannte Simulationsexperiment von Miller und Urey (Abb. 2A).
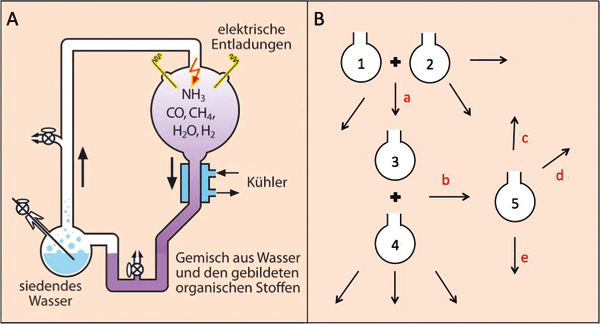 |
| Abb. 2: Prinzip eines klassischen Eintopfexperiments (A) (hier: Miller/Urey-Simulationsexperiment) im Vergleich zum Konzept von Patel et. al (2015) (B). Für den experimentellen Ansatz von Patel et al. (2015) stellt die Abbildung eine starke schematische Vereinfachung dar: Jeder einzelne Ansatz (1-5) besteht aus einem komplexen experimentellen Aufbau, und zwischen den Versuchen müssen die gewünschten Produkte durch unterschiedliche komplexe technische Verfahren (z. B. Säulenchromatographie) aufgereinigt werden. Die unbeschrifteten Pfeile deuten die Existenz weiterer Nebenprodukte an. |
|
Sutherland ist sich dieses Problems bewusst. Daher beschreibt er die Probleme der bekannten Eintopfszenarien. Als Problem bekannter präbiotischer Eintopfszenarien beschreibt Sutherland die „geringe Ausbeute biologisch relevanter Produkte aufgrund unbegünstigter Reaktionen und/oder Reaktionssequenzen“ (übersetzt nach Patel et al. 2015). Dabei stellt sich das Problem, dass Sutherland immer nur die gewünschten Chemikalien zusammenmischt und daher potentielle begünstigtere Reaktionen nicht entdeckt, die zu anderen, in diesem Zusammenhang ungewünschten Produkten führen. Alle beschriebenen Produkte würden auch Nebenreaktionen zu unerwünschten Produkten eingehen, wenn andere Stoffe vorhanden wären. Häufig können diese im Vergleich zu den beschriebenen Reaktivitäten bevorzugt sein. Außerdem wurden alle Reaktionsparameter bei den einzelnen Reaktionen verändert (unterschiedliche Katalysatoren, Temperaturen, pH-Werte, Lösemittel [z. B. DMF – ein komplexer Stoff, der selbst erst entstehen müsste], Reaktionsdauern). Sutherland gibt an, dass er sich vom Wunsch eines Eintopfsyntheseszenarios verabschieden musste. Verabschiedet er sich damit auch von präbiotisch relevanter Forschung? Es ist notwendig, sich dieser Frage zu stellen.
|
| Sutherland gibt an, dass er sich vom Wunsch eines Eintopfsyntheseszenarios verabschieden musste. Verabschiedet er sich damit auch von präbiotisch relevanter Forschung? |
|
|
Sutherland schlägt in seiner Arbeit neue Denkansätze vor. Dabei bedient er sich moderner Methoden aus der organischen Chemie (Flow Chemistry) und schlägt vor, dass unterschiedliche chemische Bedingungen in Flussströmen zur Bildung unterschiedlicher Substanzen führten, die dann an einem Knotenpunkt aufeinandertreffen und miteinander reagieren. Dieses Szenario ist ein interessanter Ansatzpunkt, aber sowohl die geochemischen Voraussetzungen wie zum Beispiel hohe Cyanid-Konzentrationen als auch die Wahrscheinlichkeit jeweils günstigerer Bedingungen in Flussströmen für die Bildung vorgeschlagener Substanzen ist sehr fragwürdig und wird auch sehr skeptisch aufgenommen. So sagt z. B. Nick Lane (University College London) in einem Kommentar: „Ich halte den geochemischen Kontext für unwahrscheinlich. Es braucht sehr hohe Cyanid-Konzentrationen und es gibt keine Belege, die dies stützen“ (übersetzt nach Extance 2015).
Dass Sutherlands jüngste Forschung einen entscheidenden Fortschritt darstellt, muss also in vielerlei Hinsicht sehr kritisch hinterfragt werden. Insbesondere der gewählte Grundansatz erscheint fragwürdig und kann kaum entscheidende Erkenntnisfortschritte im Vergleich zu Eintopfsynthesen à la Miller mit sich zu bringen. Man könnte diesen Ansatz auch für das Simulationsexperiment von Miller wählen und gewünschte Bausteine isolieren und dann gezielt unter anderen Bedingungen zur Reaktion bringen. Dies wird vermutlich nicht gemacht, weil man sich der geringen präbiotischen Relevanz eines solchen Experiments bewusst ist.
|
