 |
25 Jahre Evolution in vitro
von Daniel Vedder
Studium Integrale Journal
21. Jahrgang / Heft 1 - Mai 2014
Seite 36 - 38
|
 |
Zusammenfassung: Ein Langzeit-Evolutionsexperiment mit Bakterien erreicht demnächst 60.000 Generationen. In den letzten Jahren wurden bereits interessante Ergebnisse publiziert, etwa zum Auftauchen aerober Citratnutzung in E. coli oder über die Rolle von Hypermutatoren. Nun ist eine neue Studie erschienen, die nahelegt, dass die evolutionäre Adaptation auch bei gleichbleibenden Bedingungen nicht aufhört. Ob neue Strukturen evolutionär entstehen können, wurde bisher jedoch nicht gezeigt. | |
|
Das Langzeit-Evolutionsexperiment
Eines der größten Hindernisse der experimentellen Evolutionsbiologie sind die sehr langen Zeitspannen, mit denen gerechnet werden muss. Zwar lassen sich im Freiland Populationen unter mehr oder minder natürlichen Bedingungen beobachten, doch immer nur für relativ wenige Generationen. Die Generationszeit der Studienorganismen erweist sich in der Praxis als limitierender Faktor, weswegen nur wenige Experimente länger laufen als 100 oder sogar 1000 Generationen. Ein Freiland-Experiment mit Anolis-Eidechsen etwa umfasste 15 Generationen in ebenso vielen Jahren (vgl. Vedder 2012). Wenn von einer Erdgeschichte von mehreren Milliarden Jahren ausgegangen wird, leidet natürlich die Aussagekraft der Ergebnisse, die aus Experimenten mit so wenigen Generationen gewonnen werden. Um diese Problematik zu umgehen, werden in Evolutionsexperimenten bevorzugt Modellorganismen mit einer möglichst kurzen Generationsdauer eingesetzt, wie die Taufliege Drosophila melanogaster oder der Fadenwurm Caenorhabditis elegans, deren Reproduktionszeit in Tagen oder Wochen statt in Monaten oder Jahren gemessen wird. Dadurch lassen sich in relativ kurzer Zeit viele Generationen beobachten und so aussagekräftigere Ergebnisse erzielen.
Ein anderer beliebter Modellorganismus für solche Experimente ist das Darmbakterium Escherichia coli. Es bietet zwar als Einzeller nur begrenzte Forschungsmöglichkeiten, durchläuft dafür aber an einem einzigen Tag sechs bis sieben Generationen. Aus diesem Grund entschied sich der amerikanische Evolutionsbiologe Richard Lenski, mit E. coli als Studienorganismus zu arbeiten, als er sein Langzeit-Evolutionsexperiment (Long-Term Evolution Experiment, LTEE) begann. Für seine Versuche startete er mit 12 verschiedenen Bakterienpopulationen („Stämme“) in einem Glucose-Nährmedium. Jeden Tag werden aus jedem Stamm einige Bakterien nach dem Zufallsprinzip entnommen und in frisches Medium gegeben, in welchem sie sich weiterentwickeln können. Regelmäßig eingefrorene Proben ergeben eine Art „Fossilbericht“, der benutzt werden kann, um Bakterien unterschiedlicher Generationen miteinander bezüglich evolutionärer Fitness oder sonstiger Eigenschaften zu vergleichen und ggf. auch ausgewählte Etappen zu wiederholen.
 |
| Abb. 1: Die 12 evolvierenden E. coli-Populationen am 25. Juni 2008. (Brian Baer und Neerja Hajela, Creative Commons Attribution-Share Alike 1.0 Generic license) |
|
Was 1988 als kleines Projekt begann, wurde bald weithin bekannt und erregte immer mehr Aufsehen. Anlässlich des 25-jährigen Jubiläums publizierte die Wissenschaftszeitschrift „Science“ letztes Jahr sogar einen eigenen Bericht darüber (Pennisi 2013). So sind die bis jetzt erreichten ca. 60.000 Generationen rein rechnerisch vergleichbar mit einer Million Jahren Evolution des Menschen, womit es das derzeit mit Abstand am längsten laufende Evolutionsexperiment ist.
Das LTEE hat bisher einige interessante Ergebnisse hervorgebracht: Beispielsweise sind sechs der Stämme zu so genannten Hypermutatoren geworden, d. h. ihre Mutationsrate ist um mehrere Größenordnungen gestiegen. Unerwartet war, dass sich die Stämme trotzdem insofern ziemlich parallel entwickelten, als der Fitnessgraph jedes Stammes dieselbe Form aufwies (Abb. 2). Das bekannteste und bedeutendste Ergebnis ist jedoch das Auftauchen eines Citrattransports in E. coli, das für viel Furore sorgte. Die Charakterisierung von E. coli zeigt, dass die Mikroben unter aeroben Bedingungen Citrat nicht in die Zelle transportieren können, und daher unter solchen Bedingungen das Citrat trotz des verfügbaren Zitronensäurezyklus nicht als Kohlenstoffquelle nutzen kann. Blount et al. (2008) hatten berichtet, dass in einem der 12 E. coli-Stämme diese Funktion über mehrere Mutationen erlangt worden ist, dies war als „evolutionäre Neuheit“ beschrieben worden. Bei genaueren Untersuchungen stellte sich jedoch heraus, dass die vermeintliche „key innovation“ lediglich auf Veränderungen in der Genregulation zurückzuführen ist, also keineswegs die Entstehung einer neuen Funktion vorliegt (vgl. Binder 2012).
Zwischendurch experimentierte Lenski auch mit digitalen Organismen (kleine selbst-replizierende Computerprogamme), die eine noch viel kürzere Generationszeit aufweisen und außerdem einfacher zu handhaben und untersuchen sind als Bakterien. Die Relevanz dieser Ergebnisse für reale biologische Systeme ist aber zweifelhaft (vgl. Bertsch & Waldminghaus 2005).
|
|
|
Unendliche Evolution?
Neueste Ergebnisse aus dem Lenski-Labor beziehen sich auf den Gesamt-Fitnessgewinn der Populationen, der über 50.000 Generationen des Experiments hinweg gemessen wurde (Wiser et al. 2013). Im Schnitt waren die Bakterien in dem verwendeten Nährmedium 1,7x fitter als die ursprüngliche Gründerpopulation, gemessen an ihrer relativen Wachstumsrate. In den frühen Phasen des LTEE stieg die Fitness am schnellsten, danach flachte die Kurve ab, stieg aber immer weiter. Nach 10.000 Generationen hatten die Forscher vorausgesagt, dass es irgendwann keine vorteilhaften Mutationen für die gleichbleibenden Bedingungen geben und die Fitness somit nicht weiter gesteigert würde (Lenski & Travisano 1994). Es gibt jedoch immer noch keine Anzeichen für ein Fitnessplateau. Im Gegenteil, mathematische Beschreibungen der Fitnesskurve über die Zeit legen ein Modell nahe, dessen Steigung nie Null wird, d. h. dass die Fitness also immer größer wird. Interessanterweise haben die oben erwähnten Hypermutatoren fast durchgehend eine höhere Fitness als Stämme mit normalen Mutationsraten (Abb. 2).
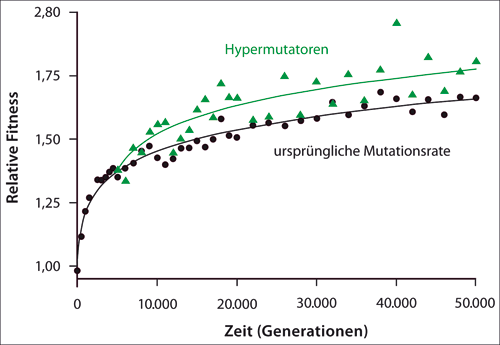 |
| Abb. 2: Fitnesszunahme der 12 Stämme im Laufe von 50.000 Generationen. Datenpunkte von Hypermutatoren-Stämmen sind als Dreiecke eingetragen, ihre höhere Fitness ist deutlich erkennbar. Die Linien stellen das jeweils am besten passende mathematische Modell für Hyper- bzw. Nichtmutatoren dar. (Erklärung im Text; nach Wiser et al. 2013) |
|
|
|
Diskussion
Dass die Fitness scheinbar unendlich steigt, ist ein unerwartetes Ergebnis. Nach dem gängigen Modell der Fitnesslandschaft entwickeln sich Populationen unter gleichbleibenden Bedingungen in Richtung einer „Bergspitze“, eines Maximums. Ist die Spitze der Landschaft erreicht, kann die Anpassung mangels Möglichkeiten nicht weiter verbessert werden. Natürlich ist es noch möglich, dass das LTEE diesen Zustand trotzdem irgendwann erreichen wird. Pennisi zitiert jedoch Lenski mit folgenden Worten: „Das Konzept einer Fitness-Spitze ist schwerer zu greifen als ich erwartet hatte. Ich halte es für gut möglich, dass die Fitness sich noch eine Million Jahre lang weiter steigern wird“ (Pennisi 2013, übersetzt vom Autor). Auch aus der Sicht der Schöpfungsforschung ist das durchaus überraschend. Lönnigs Regel der rekurrenten Variation besagt, dass einem Grundtyp nur endlich viele verschiedene Mutationen zur Verfügung stehen (vgl. Lönnig 2005). Daher würde man auch hier schlussendlich ein Fitnessmaximum erwarten. Warum dies (noch) nicht eingetroffen ist, bleibt offen. Es muss allerdings auch bedacht werden, dass Fitnesssteigerung nicht gleichbedeutend ist mit der Entwicklung evolutionärer Neuheiten.
Außerdem ist bei der Bewertung der Ergebnisse natürlich zu berücksichtigen, dass dies nur die Daten aus einem einzigen Großexperiment sind. Weitere Ergebnisse könnten das Bild stark verändern. Sollte das aber nicht der Fall sein, müssen einige theoretische Modelle modifiziert oder gar verworfen werden.
 |
Warum noch kein Fitnessmaximum
erreicht wurde, ist unklar. |
 |
|
Die hervorgehobene Rolle der Hypermutatoren ist auch eine besondere Betrachtung wert. Da die meisten Mutationen der Fitness abträglich sind, war man früher davon ausgegangen, dass die natürliche Selektion die Mutationsrate so gering wie möglich halten würde. Daher mussten, als 1997 im LTEE die Hypermutatoren beschrieben wurden, die sich gegen Nicht-Mutatoren durchsetzen konnten, neue evolutionsbiologische Modelle zum Einfluss der Mutationsrate entwickelt werden (Sniegowski et al. 1997). Der Biochemiker Michael Behe macht außerdem darauf aufmerksam, dass einige der Hypermutatoren-Stämme ihren Phänotyp durch die Ausschaltung zweier Enzyme erlangten. Somit wäre diese evolutionär vorteilhafte Entwicklung eine Rückentwicklung – „ein schlechtes Omen für jegliche Theorie der Evolution, die sich allein auf blinde, ungerichtete Prozesse verlässt“, wie er sich ausdrückt (Behe 2013, übersetzt vom Autor). Ob zudem Hypermutatoren makroevolutive Änderungen erleichtern oder ermöglichen, ist offen und durch die bisher vorliegenden Befunde aus dem LTEE nicht entscheidbar.
Das LTEE bleibt auch nach 25 Jahren ein faszinierendes Experiment, das viele wertvolle Daten liefert. Vielleicht führen einige davon dazu, dass momentan anerkannte Hypothesen revidiert werden müssen – das ist der Gang der Wissenschaft. Ob nun die natürliche Selektion tatsächlich unendlich weiterläuft, wie Lenski vermutet, oder ob sie doch in einem Plateau mündet müssen zukünftige Untersuchungen zeigen.
|
Literatur
- Bertsch E & Waldminghaus T (2005)
- Evolution virtueller Lebewesen. Stud. Integr. J. 12, 34-35.
- Binder H (2012)
- Von der Citrat-Verwertung zur Entstehung des Auges? Genesisnet News vom 23.10.2012, http://www.genesisnet.info/schoepfung_evolution/n187.php
- Behe M (2013)
- „Lenski‘s Long-Term Evolution Experiment: 25 Years and Counting”, Evolution News http://www.evolutionnews.org/2013/11/richard_lenskis079401.html
- Blount ZD, Borland CZ & Lenski RE (2008)
- Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli. Proc. Natl. Acad. Sci. 105, 7899-7906.
- Lenski RE (2011)
- Evolution in Action – a 50,000 Generation Salute to Charles Darwin. Microbe 6, 30-33.
- Lenski RE & Travisano M (1994)
- Dynamics of adaptation and diversification: a 10,000-generation experiment with bacterial populations. Proc. Natl. Acad. Sci. 91, 6808-6814.
- Lönnig WE (2005)
- Mutation Breeding, Evolution and the Law of Recurrent Variation. Recent Research Developments in Genetics and Breeding 2, 45-70.
- Pennisi E (2013)
- The man who bottled evolution. Science 342, 790-793.
- Sniegowski PD, Gerris PJ & Lenski RE (1997)
- Evolution of high mutation rates in experimental populations of E. coli. Nature 387, 703-705.
- Vedder D (2012)
- Gründereffekt bei Eidechsen: ein Freilandexperiment auf Inseln. Stud. Integr. J. 19, 107-109.
- Wiser MJ, Ribeck N & Lenski R (2013)
- Long-term dynamics of adaption in asexual populations. Science 342, 1364-1367.
-
|
|  |
