 |
„Homo“ Habiliswar kein Mensch
Kluft zwischen fossilen Menschen und Menschenaffen größer geworden
von Michael Brandt
Studium Integrale Journal
19. Jahrgang / Heft 1 - Mai 2012
Seite 4 - 11
|
 |
Zusammenfassung: „Homo“ habilis/rudolfensis gilt als evolutionär vermittelnde Fossilgruppe zwischen den Australopithecinen und dem frühesten echten Menschen (Homo erectus). Doch es gibt gute Gründe, diese Fossilgruppe aus Homo auszugliedern. Damit wird die Lücke zwischen ausgestorbenen menschenaffenartigen Primaten und dem Menschen größer.
| |
|
Einleitung
1999 berichtete Hartwig-Scherer in dieser Zeitschrift unter der Überschrift „Homo“ habilis ab jetzt kein Mensch mehr über eine bemerkenswerte Umklassifizierung von Homo habilis und Homo rudolfensis. Wood & Collard (1999) hatten beide Homininen*-Arten aus der Gattung Homo – Mensch – herausgenommen und zur ausgestorbenen Gattung Australopithecus gestellt. Die Australopithecinen werden im evolutionären Stammbaum als „Vormenschen“ gedeutet. Plausibler scheint jedoch die Annahme, des es spezialisierte Menschenaffen gewesen sind. Obwohl Bernard Wood, einer der Päpste der Paläanthropologie, gute Gründe für die Reklassifizierung angeführt hatte, folgte ihm kaum jemand auf diesem Weg. Beide Arten wurden in den allermeisten paläanthropologischen Facharbeiten weiterhin zu Homo gestellt und als primitivste Formen des Menschen interpretiert.
Die Folgenlosigkeit der Arbeit von Wood & Collard (1999) war nicht unbedingt zu erwarten gewesen. Man kann heute nur vermuten, dass die nicht unerheblichen Konsequenzen im Evolutionsmodell des Menschen der Hauptgrund dafür sind. Denn beide Arten werden als phylogenetisch vermittelnde Homininen zwischen den recht großaffenähnlichen Australopithecinen und dem frühesten unzweifelhaft echten Menschen Homo erectus angesehen (Abb. 1, 2). Sie schienen also gut als Bindeglieder zu passen, die unmittelbar zum Menschen führen.
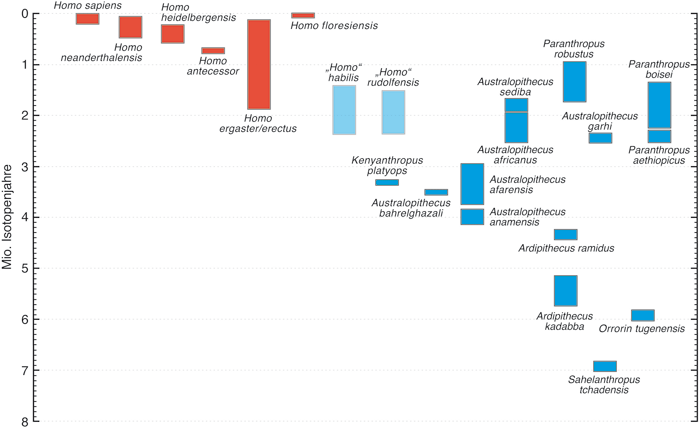 |
| Abb. 1: Die fossilen Homininen-Arten und ihr zeitliches Auftreten. Nach Wood & Baker (2011), geringfügig verändert. |
|
Seit 1999 sind weitere überzeugende Untersuchungsergebnisse publiziert worden, die auf eine Neuklassifizierung drängen. Im Folgenden werden diese Befunde dargestellt. Wegen der notwendigen taxonomischen Revision wird der Gattungsname Homo der beiden Arten habilis und rudolfensis in Anführungszeichen gesetzt.
|
„Homo“ habilis/rudolfensis wird seit Jahrzehnten als evolutionär vermittelnde Fossilgruppe zwischen den Australopithecinen und dem frühesten echten Menschen (Homo erectus) verstanden. Neuere Untersuchungen zeigen jedoch, dass „Homo“ habilis ein Belastungsverhältnis von Vorder- zu Hinterextremitäten ähnlich dem der Schimpansen hatte und sich wie die Australopithecinen ernährte. Bei Homo erectus war dagegen die Ernährung und das Extremitätenbelastungsmuster menschlich modern. Eine notwendige Neurekonstruktion des berühmten Schädels KNM-ER 1470 rückt „Homo“ rudolfensis noch näher als zuvor an die robusten Australopithecinen. KNM-ER 1470 zeigt außerdem verblüffende Ähnlichkeiten mit dem neu entdeckten Homininen Kenyanthropus platyops. Es gibt damit gute Gründe, die beiden Arten habilis und rudolfensis aus Homo auszugliedern und zu den recht großaffenähnlichen Gattungen Australopithecus bzw. Kenyanthropus zu stellen.
Mit dieser Rückstufung erscheint die Lücke zwischen den mehr großaffenähnlichen frühen Homininen einerseits und den verschiedenen Formen des echten Menschen andererseits deutlicher.
Aus schöpfungstheoretischer Sicht lassen sich Homo erectus und die späteren Homo-Formen dem Grundtyp Mensch zuordnen. Die mehr großaffenähnlichen frühen Homininen können dagegen zu anderen Grundtypen ohne historisch-verwandtschaftliche Beziehung zum Menschen gestellt werden. |
|
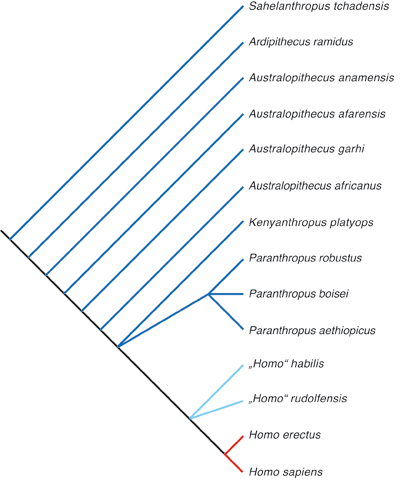 |
| Abb. 2: Kladogramm, das eine repräsentative Hypothese über die Verwandtschaftsbeziehung der frühen Homininen aufzeigt. Nach Wood & Baker (2011), geringfügig verändert. |
|
|
|
Befunde und Interpretationen im vergangenen Jahrhundert
Ende der 1950er und Anfang der 1960er Jahre wurden aus der Olduvai-Schlucht in Tansania Skelettfragmente und Steinwerkzeuge geborgen, die als Indizien für eine neue Homininen-Art angesehen wurden und den Namen „Mensch“ rechtfertigten. Das neue Taxon* „Homo“ habilis wurde durch Leakey, Tobias und Napier 1964 eingeführt. Dieses Wesen hatte ein etwas größeres Schädelvolumen als die Australopithecinen. Ihm wurden eine menschlich-zweibeinige Fortbewegung und die Fähigkeit zur Werkzeugherstellung zugeschrieben.
Die Zuordnung von habilis zur Gattung Homo war von Anfang an umstritten. Insbesondere war das auf ca. 670-680 cm3 geschätzte Schädelvolumen kleiner als die zu dieser Zeit (ohne besonders überzeugende Gründe) allgemein akzeptierte minimale Gehirngröße für Homo von 700 bis 800 cm3 (Wood & Baker 2011). So gab es damals schon Forscher, die das Olduvai-Material alternativ Australopithecus zuordneten. Die Zuordnung zur Gattung Homo setzte sich durch, obwohl zahlreiche spätere Untersuchungen deutliche Ähnlichkeiten mit den Australopithecinen aufzeigten. So weist „Homo“ habilis Kletteranpassungen und Merkmale einer vom Menschen verschiedenen Bipedie (Zweibeinigkeit) auf (Übersicht bei Brandt 1995).
|
Bipedie: Zweibeinigkeit. Holotyp: Dasjenige Exemplar, welches in der Originalbeschreibung als der „Typus“ festgelegt wird. Homininen (Menschenähnliche): Damit werden alle fossilen und lebenden Menschenformen einschließlich ihrer im Evolutionsmodell vermuteten Vorläufer bezeichnet. Vorläufer-Arten: Ardipithecus ramidus, Ar. kadabba, Australopithecus afarensis, Au. africanus, Au. anamensis, Au. bahrelghazali, Au. garhi, Au. („Homo“) habilis, Au. sediba, Paranthropus aethiopicus, P. boisei, P. robustus, Kenyanthropus platyops, K. („Homo“) rudolfensis, Orrorin tugenensis, Sahelanthropus tchadensis (siehe Abb. 1). Humerofemoralindex: Längenverhältnis von Oberarm zu Oberschenkel; der Index wird bestimmt, indem die Länge des Oberarmknochens durch die Länge des Oberschenkelknochens dividiert und mit 100 multipliziert wird. Morphologie: Wissenschaft vom Bau und der Gestalt des Körpers der Lebewesen und seiner Organe. postcranial: dem Skelett mit Ausnahme des Schädels zugehörig. Taxon: eine als systematische Einheit erkannte Gruppe von Lebewesen.
|
Nach Aufstellung der Art „Homo“ habilis wurden ab 1972 Fossilien (Schädel, Zähne) in Koobi Fora, Kenia, entdeckt, die man ebenfalls dem frühen Homo zuordnete. Das neue Material unterschied sich wie das von Olduvai von Homo erectus.
Sehr bald zeigte sich aber, dass das gesamte zum frühen Homo gestellte Material die Variationsbreite eines einzelnen Taxons überschreitet. Während einige Forscher jedoch für die Beibehaltung eines einzigen Taxons mit nun erweitertem Fossilmaterial („Homo“ habilis sensu lato) plädierten, unterstützten andere die Aufstellung eines zweiten Taxons, „Homo“ rudolfensis, mit dem Schädel KNM-ER (Kenya National Museums – East Rudolf) 1470 als wichtigstem Fundstück. KNM-ER 1470 weist ein Paranthropus-ähnliches flaches, hohes und weites Mittelgesicht und einen mehr Homo-ähnlich großen, zunächst auf 750-800 cm3 geschätzten Hirnschädel auf. Dagegen sind die Schädel von „Homo“ habilis sensu stricto (OH [Olduvai Hominid] 13, KNM-ER 1813) im oberen Gesichtsbereich am breitesten und das absolute Schädelvolumen (z. B. OH 7 ca. 670 cm3) kleiner (Wood & Baker 2011).
Die meisten Forscher maßen dem Schädelvolumen eine größere Bedeutung als den Paranthropus-ähnlichen Gesichtsmerkmalen für die taxonomische Zuordnung bei und stellten KNM-ER 1470 deshalb zu Homo (Wood 2009). Unter den frühen Bearbeitern mahnte aber Walker (1976) zur Vorsicht in Bezug auf diese taxonomische Klassifizierung.
1999 stellten Wood & Collard eine Diagnoseliste auf (Tab. 1), anhand derer Funde nach ihrer möglichen Zugehörigkeit zur Gattung Homo beurteilt werden können. Um Homo zugeordnet zu werden, müssen Funde in ihrer Organisationshöhe (Anpassung) dem modernen Menschen näher stehen als irgendeiner australopithecinen Form. Keine der geforderten deutlichen Abgrenzungen zu den australopithecinen Formen trifft jedoch nach ihrer Analyse für habilis und rudolfensis zu. Beide Arten gehören deshalb nicht zur Gattung Homo. Wood & Collard (1999) stellten sie vorläufig zur Gattung Australopithecus.
|
Nach Wood & Collard (1999) muss gezeigt werden, dass
1. die Form näher mit H. sapiens als mit einer der Australopithecinen verwandt ist,
2. ihr Körpergewicht mehr H. sapiens als einem der Australopithecinen ähnelt,
3. ihre Körperproportionen mehr H. sapiens als einem der Australopithecinen ähneln,
4. sie einen zweibeinigen aufrechten Gang ohne wesentliche Kletterfähigkeit besaßen,
5. die relative Zahn- und Körpergröße mehr H. sapiens als einem Australopithecinen ähnelt,
6. sie eine für den Menschen typische verlängerte Kindheitsentwicklung zeigen.
|
|
| Tab. 1: Diagnoseliste zur Identifikation von Funden zur möglichen Aufnahme in die Gattung Homo. Aus Hartwig-Scherer (1999). |
|
Neue Untersuchungsergebnisse erhärten die Notwendigkeit einer Reklassifizierung von habilis und rudolfensis.
|
|
Neuere Untersuchungsergebnisse
Die Fortbewegung von „Homo“ habilis
Die im Folgenden aufgeführten Untersuchungen und Ergebnisse zur Fortbewegung von „Homo“ habilis sind ausführlicher im Kasten „Neuere Untersuchungsergebnisse“ dargestellt.
Neuere Untersuchungen bestätigen, dass der von vielen Paläanthropologen zu „Homo“ habilis gestellte Teilfuß OH 8 (Leakey et al. 1964, Susman 2008) sich in seinem Gesamtmerkmalskomplex deutlich vom Menschen unterscheidet.
Die Teilhand OH 7 wurde zusammen mit Resten des Schädels und Zähnen entdeckt und gehört zum Holotyp* von „Homo“ habilis (Leakey et al. 1964). OH 7 besitzt großaffenähnliche Merkmale mit Anpassungen an das Klettern (Übersicht bei Brandt 1995). Eine Untersuchung des Trapeziums (Großes Vielecksbein, Handwurzelknochen) von OH 7 erbrachte das erstaunliche Ergebnis, dass dieser Handwurzelknochen primitiver (großaffenähnlicher) ist als das Trapezium von Australopithecus afarensis.
Der Teilfuß OH 8 und auch die Teilhand OH 7 werden von einigen Forschern nicht zu „Homo“ habilis, sondern zu den robusten Australopithecinen gestellt.
Unbestritten „Homo“ habilis zuzuordnen ist das mit Resten von Schädel und Zähnen assoziierte Teilskelett OH 62 (Johanson et al. 1987) und wahrscheinlich auch das Teilskelett KNM-ER 3735 (Leakey et al. 1989).
Johanson et al. (1987) und Hartwig-Scherer & Martin (1991) kamen auf der Basis von Längenmessungen der Gliedmaßen zu dem Ergebnis, dass OH 62 großaffenähnlichere Körperproportionen als A.L. 288-1 (Au. afarensis) besitzt. Diese Ergebnisse überraschten, weil „H.“ habilis im Bereich des Schädels und der Zähne deutlich menschenähnlicher als Au. afarensis ist.
Gegenüber dieser Schlussfolgerung erhob sich Kritik, wobei im Mittelpunkt die Längenbestimmung des nicht vollständig erhaltenen Femurs (Oberschenkelknochen) von OH 62 stand. Zunächst konnten nachfolgende Studien die Ergebnisse von Johanson et al. (1987) und Hartwig-Scherer & Martin (1991) nicht bestätigen. Aber damit war nicht das letzte Wort gesprochen.
|
Morphologie und Taxonomie von OH 7 (Teilhand) und OH 8 (Teilfuß)
Nach Kidd (1999) zeigt OH 8 eine mosaikartige Morphologie* und Funktion. Der Talus (Sprungbein) ist dem des Orang Utan sehr ähnlich. Das Naviculare (Kahnbein) ist ebenfalls eindeutig großaffenähnlich mit der größten Nähe zum Gorilla. Das Cuboid (Würfelbein) ist dem des Menschen am ähnlichsten. Der Calcaneus (Fersenbein) ist wegen seiner starken Fragmentierung nur eingeschränkt beurteilbar. Mit seiner einmaligen Morphologie unterscheidet er sich von dem der Großaffen und des Menschen.
Der Talus von OH 8 besitzt nach Gebo & Schwartz (2006) ähnliche Merkmale wie der Paranthropus robustus zugeordnete Talus TM (Transvaal Museum) 1517. OH 8 wird deshalb von beiden Autoren auch zu diesem Taxon und nicht zu „Homo“ habilis gestellt.
Nach Wood & Constantino (2007) können die „Homo“ habilis zugeordneten postcranialen* Skelettelemente OH 8 (Teilfuß), OH 10 (Großzehenendglied) und OH 35 (Unterschenkelknochen) zu Paranthropus boisei gestellt werden.
Aber auch die mit craniodentalen Resten assoziierte OH 7-Teilhand als Bestandteil des Holotyps von wird bestritten und eine Zuordnung zu Paranthropus boisei diskutiert, denn sie zeigt Ähnlichkeiten mit diesem Taxon (Moyà-Solà et al. 2008).„Homo“ habilis
In Übereinstimmung mit diesen Analysen steht das Untersuchungsergebnis von Trocheri et al. (2003), nach dem das Trapezium (Großes Vielecksbein, Handwurzelknochen) von OH 7 primitiver (großaffenähnlicher) ist als das Trapezium von Australopithecus afarensis (A.L. [Afar Locality] 333-80). Falls dieser Knochen Teil der OH 7-Hand ist, die für die Herstellung und Nutzung der Steinwerkzeuge von Olduvai verantwortlich ist, dann tat sie es in einer Weise – so schließen die Autoren –, für die es keine moderne Analogie gibt. Für Trocheri et al. (2003) verbleiben noch zwei weitere Interpretationsmöglichkeiten: Der Handwurzelknochen ist entweder nicht repräsentativ für„Homo“ habilis oder er gehört zu einer anderen Primaten- oder Homininen-Art.
Strittige Ergebnisse zu den Körperproportionen von OH 62 („H.“ habilis) und A.L. 288-1 (Au. afarensis)
OH 62 weist nach Johanson et al. (1987) mit 95% einen schimpansenähnlicheren Humerofemoralindex* und damit großaffenähnlichere Körperproportionen als A.L. 288-1 (Au. afarensis) mit einem Index von 85% auf.
Hartwig-Scherer & Martin (1991) kamen auf der Basis umfangreicher Vergleichsmessungen zwischen Vorder- und Hinterextremitäten ebenfalls zu dem Ergebnis, dass OH 62 mit seinen Körperproportionen den Afrikanischen Großaffen näher steht als A.L. 288-1 (Au. afarensis).
Nach Korey et al. (1990) ist die Unsicherheit bei der Bestimmung der Femurlänge bei diesem Teilskelett jedoch so groß, dass OH 62 nicht sicher von A.L. 288-1 getrennt werden kann. Der Femur von OH 62 liegt aber näher am Schimpansen als am modernen Menschen (Abb. 3).
Richmond et al. (2002) fanden keinen statistisch signifikanten Unterschied im Größenverhältnis von Humerus (Oberarmknochen) zu Femur (Oberschenkelknochen) bei OH 62 und A.L. 288-1.
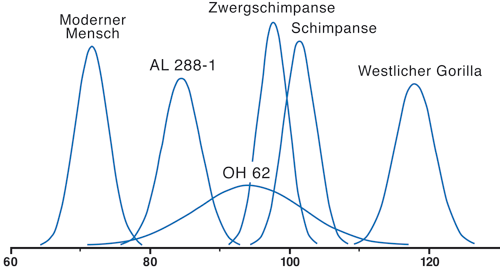 |
| Abb. 3: Stichprobennormalverteilung des Humerofemoralindex von vier modernen Hominoiden (moderner Mensch, Schimpanse, Zwergschimpanse, Westlicher Gorilla) und extrapoliert für A.L. 288-1 (Australopithcus afarensis) sowie mit einer ähnlichen Fehlerverteilung für OH 62 („Homo“ habilis) bestimmt. Nach Korey (1990). |
|
Reno et al. (2005) vermuten, dass alle durchgeführten Bestimmungen der Länge des Femurs von OH 62 nicht überzeugend seien und der humerofemorale Index von OH 62 deshalb letztlich unbekannt bleibe.
Haeusler & McHenry (2004, 2007) schlussfolgern aus ihren Untersuchungen, dass die Vorder- zu Hinterextremitätenproportion bei OH 62, KNM-ER 3735 (beide „H.“ habilis) und A.L. 288-1 (Au. afarensis) in den menschenähnlichen Variationsbereich fallen, wobei OH 62 aufgrund seiner geringen Größe aber auch eine Überlappung mit den Schimpansen aufweist. Haeusler & McHenry (2007) kommen aber auch zu dem Ergebnis, dass „Homo“ habilis schimpansenähnlich lange Unterarme, Australopithecus afarensis dagegen menschenähnlich kurze Unterarme besaß.
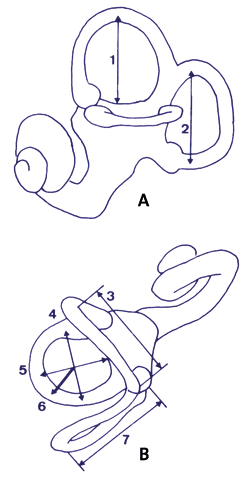 |
| Abb. 5: Das knöcherne Labyrinth mit Ansicht von (A) lateral und (B) superior (nach Spoor et al. 1994). Die drei Bogengänge stehen senkrecht aufeinander. Die relative Größe der Bogengänge ermöglicht Rückschlüsse auf die Körperhaltung. Abkürzungen: (1) Höhe des Ductus semicircularis anterior, (2) Weite des Ductus semicircularis posterior, (3) Weite des Ductus semicircularis anterior, (4) Weite des Ductus semicircularis lateralis, (5) Höhe des Ductus semicircularis lateralis, (6) Radius des Ductus semicircularis lateralis, (7) Höhe des Ductus semicircularis posterior. |
|
Gliedmaßenbelastung von OH 62 („H.“ habilis) ähnlich der von Schimpansen
2009 publizierte Ruff die vielleicht eindrucksvollste Studie zu diesem Thema, bei der die Fehlergröße bei der Femurlängenbestimmung keine Rolle spielt. Ruff (2009) benutzte Messdaten von nicht rekonstruierten fossilen Knochen. Er ermittelte die Knochenquerschnittsstärke an zwei Stellen des Femurs und des Humerus von OH 62 und verglich diese mit denen vom Schimpansen, modernen Menschen und Homo erectus (KNM-WT [Kenya National Museums-West Turkana] 15000, KNM-ER 1808). Die Kombinationen der Querschnittsstellen weisen alle ein Femur-Humerus-Stärkenverhältnis unterhalb des 95%igen Vertrauensintervalls der Werte des modernen Menschen und mit einer Ausnahme im 95%igen Vertrauensintervall der Werte des Schimpansen auf. Die beiden Homo erectus-Teilskelette zeigen ein Femur-Humerus-Stärkenverhältnis im oder oberhalb des Variationsbereichs des modernen Menschen (Abb. 4). Carlos et al. (2010) konnten die Ergebnisse von Ruff (2009) bestätigen.
Das verschiedene Lastverhältnis von Vorder- zu Hinterextremitäten bei „Homo“ habilis und Homo erectus weist auf deutliche Unterschiede in der Fortbewegungsweise hin.
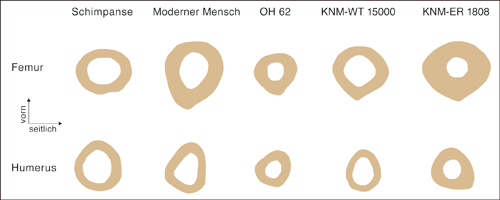 |
| Abb. 4: Repräsentative Querschnitte vom Femur (Oberschenkelknochen) und Humerus (Oberarmknochen) von Schimpansen, modernen Menschen, „Homo“ habilis (OH 62) und Homo erectus (KNM-WT 15000, KNM-ER 1808). Das Femur-Humerus-Stärkenverhältnis von „Homo“ habilis ist dem des Schimpansen und das von Homo erectus dem des modernen Menschen ähnlich. Nach Ruff (2009). |
|
Innenohr von „Homo“ habilis ähnlich dem von Meerkatzenartigen
Im Labyrinth des Innenohres befinden sich drei Bogengänge (Abb. 5), die Bestandteil des Gleichgewichtsorgans sind. Die relative Größe der Bogengänge weist auf die gewohnheitsmäßige Körperhaltung bei Primaten hin.
Homo erectus besitzt wie der moderne Mensch einen größeren vorderen und hinteren und einen kleineren seitlichen Bogengang. Dagegen sind die Proportionen der Bogengänge bei Australopithecus africanus und Australopithecus afarensis ähnlich denen der Großaffen. Daraus ist zu schließen, dass diese fossilen Homininen schlecht an einen aufrechten Gang angepasst waren. Noch überraschender war jedoch das Ergebnis für Stw (Swartkrans Witwatersrand) 53, einem „Homo“ habilis oder anderen frühen Homo-Taxon zugeordneten Schädel. Das Labyrinth von Stw 53 repräsentiert keine intermediäre Morphologie zwischen den Australopithecinen und Homo erectus, wie aufgrund der phylogenetisch postulierten Stellung zu erwarten war, sondern ähnelt mit seinem besonders großen seitlichen Bogengang am stärksten dem der großen Cercopithecoiden (Meerkatzenartige). Die Fortbewegung dieser Tieraffen erfolgt in der Regel im vierfüßigen Gang sowohl auf dem Erdboden als auch auf Bäumen.
|
Alle bisherigen Studien waren in ihrer Aussage durch die Unsicherheit der Rekonstruktion der fehlenden Fossilteile, insbesondere des fehlenden körperfernen Abschnittes des OH 62-Femurs, eingeschränkt. 2009 publizierte Ruff die vielleicht eindrucksvollste Studie zu diesem Thema, bei der diese Fehlergröße keine Rolle spielt.
 |
Bei „Homo“ habilis war die Belastung
der Vorder- zu Hinterextremitäten großaffenähnlich und das Innenohr gleicht dem von Tieraffen (Meerkatzenartige).
Die Fortbewegung von „Homo“ habilis unterschied sich deutlich von der menschlich-zweibeinigen Fortbewegung des Homo erectus. Neben Klettern in Bäumen muss
bei „Homo“ habilis auch eine vierfüßige Fortbewegung diskutiert werden. |
 |
|
uff benutzte Messdaten von nicht rekonstruierten fossilen Knochen. Er bestimmte die Querschnittsstärken der Oberschenkel- und Oberarmknochen von OH 62 und kam zu dem Ergebnis, dass „Homo“ habilis ein schimpansenähnliches Belastungsmuster der Gliedmaßen hatte, Homo erectus in diesem Merkmal dagegen modern menschlich war. Danach muss die Fortbewegung beider fossiler Formen sehr unterschiedlich gewesen sein.
15 Jahre vor der Analyse der Extremitätenknochenstärke gelangten Spoor et al. (1994) bezüglich der Fortbewegungsweise von „Homo“ habilis zu einem noch verblüffenderen Ergebnis. Die Forscher untersuchten nicht die Extremitätenknochen, sondern das Labyrinth des Innenohres. Die im Labyrinth befindlichen Bogengänge geben Aufschlüsse über die gewohnheitsmäßige Körperhaltung bei den Primaten. Während Homo erectus eine modern menschliche Labyrinthmorphologie besitzt, ist die der Australopithecinen großaffenähnlich. Überraschenderweise zeigt „Homo“ habilis ein im evolutionären Kontext noch primitiveres Labyrinth als die Australopithecinen, nämlich eines, das dem der Cercopithecoiden (Meerkatzenartigen) ähnelt. Diese Tieraffen bewegen sich im vierfüßigen Gang auf Bäumen und auf dem Erdboden. Bei „Homo“ habilis muss also auch eine vierfüßige Fortbewegung neben einer gelegentlichen nichtmenschlichen bipeden* Fortbewegung auf dem Erdboden und Klettern in Bäumen diskutiert werden.
Homo erectus bewegte sich dagegen effizient ausschließlich auf zwei Beinen in aufrechter Körperhaltung fort, denn seine im Zusammenhang mit der Fortbewegung stehenden Skelettmerkmale sind modern menschlich (z. B. Spoor et al. 1994, Übersicht bei Brandt 1995, Jungers 2009, Ruff 2009, Haeusler et al. 2011).
Zeitliches Auftreten
2007 wurde von Spoor et al. die Entdeckung von zwei bemerkenswerten fossilen Funden aus der Koobi Fora-Formation bei Ileret östlich des Turkana-Sees in Kenia bekannt gegeben. Bei dem einen Fundstück handelt es sich um eine Homo erectus-Schädelkalotte (KNM-ER 42700), bei dem anderen um einen rechten Oberkieferknochen von „Homo“ habilis (KNM-ER 42703). Die Schädelkalotte wird auf 1,55 Millionen Jahre und der Oberkieferknochen auf 1,44 Millionen Jahre datiert. KNM-ER 42700 liegt mit seinem Schädelvolumen von 691 cm3 zwar im oberen Grenzbereich der Schädelkapazität von „Homo“ habilis habilis, zeigt aber eine Morphologie*, wie sie typisch für den asiatischen Homo erectus ist. Allerdings wird diese taxonomische Zuordnung auch in Frage gestellt (Baab et al. 2008). Unabhängig von dieser taxonomischen Streitfrage bleibt das Fakt, dass „Homo“ habilis in Ostafrika nahezu eine halbe Million Isotopenjahre an der Seite von Homo erectus gelebt hat. Damit ist es nach Wood & Baker (2011) wenig wahrscheinlich (wenn auch nicht unmöglich), dass Homo erectus sich aus „Homo“ habilis entwickelt hat.
„Homo“ rudolfensis-Schädel KNM-ER 1470
Unter der Überschrift KNM-ER 1470 kein Vorfahr des Menschen – Aufstieg und Fall eines Stars haben Hartwig-Scherer & Brandt (2007) die Interpretationsgeschichte dieses hochkontrovers diskutierten Schädels nachgezeichnet. Zwei Ereignisse beeinflussten in jüngerer Zeit die Sicht auf diesen Schädel: zum einen eine erneute Schädelrekonstruktion und zum anderen die Entdeckung einer neuen Homininen-Gattung.
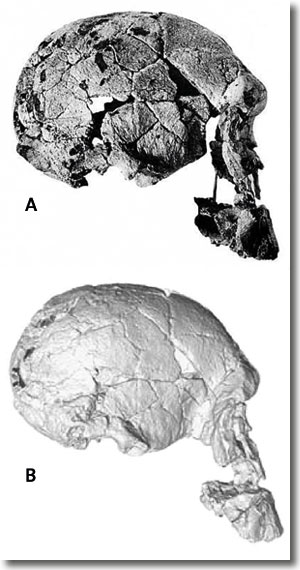 |
| Abb. 6: Rekonstruierter Schädel KNM-ER 1470 (A) nach Leakey (1972) und (B) neu virtuell nach Bromage et al. (2008). Nach der neuen Rekonstruktion zeigt KNM-ER 1470 ein stärker vorstehendes Gesicht und ein etwas kleineres Schädelvolumen. Mit diesen Merkmalen ist KNM-ER 1470 Paranthropus noch ähnlicher als zuvor. |
|
Eine neue Rekonstruktion von KNM-ER 1470 durch Bromage et al. (2007, 2008) führt zu einer etwas größeren Prognathie, d. h. zu einem stärker vorstehenden Gesicht (Abb. 6), und zu einer Reduktion des Schädelvolumens von 752 cm3 auf ca. 700 cm3 (Bromage et al. 2008). Damit rückt KNM-ER 1470 morphologisch noch weiter in die Nähe der robusten Australopithecinen als dies vorher schon der Fall war.
2001 berichteten Leakey et al. über die Entdeckung des neuen frühen Homininen Kenyanthropus platyops. Der neue Fund KNM-WT 40000 zeigt überraschenderweise eine große Ähnlichkeit mit KNM-ER 1470 (Abb. 7). Der Hauptunterschied zwischen KNM-ER 1470 und dem 1,6 Millionen Jahre älteren, neu entdeckten Fund liegt in der primitiveren Nasenmorphologie und dem deutlich kleineren Hirnschädel bei letzterem. Leakey et al. (2001) stellen deshalb KNM-ER 1470 nicht zu Australopithecus, wie es Wood & Collard (1999) vorgeschlagen hatten, sondern in die neue Gattung Kenyanthropus.
Überreste des Extremitäten- und Stammskeletts von „Homo“ rudolfensis sind bis heute nicht bekannt.
Ernährung
Die Stärke des Kauapparates von „Homo“ habilis und „Homo“ rudolfensis wurde anhand der Größe des Unterkiefers und der Kronenfläche der postcaninen Zähne (Vorderbacken- und Backenzähne) bestimmt. Wood & Aiello (1998) kamen zu dem Ergebnis, dass „Homo“ habilis und „Homo“ rudolfensis relativ größere Unterkieferknochen als Homo erectus besitzen.
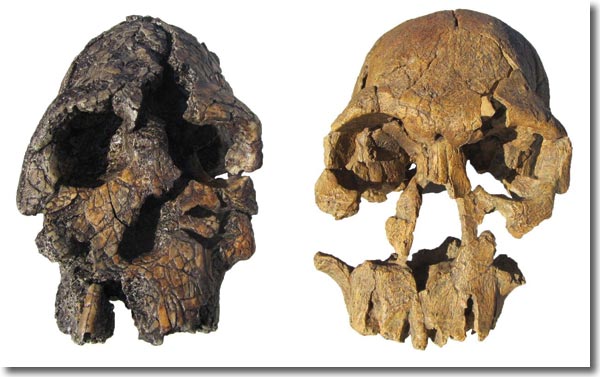 |
| Abb. 7: Kenyanthropus platyops (KNM-WT 40000; links) und „Homo“ rudolfensis (KNM-ER 1470). Der neue Hominine Kenyanthropus platyops (KNM-WT 40000) ähnelt in frappierender Weise (z. B. flaches Gesicht, Form der Augenbrauen) dem schon seit 1972 bekannten „Homo“ rudolfensis zugeordneten Schädel KNM-ER 1470. Es gibt aber auch Unterschiede, insbesondere ist der Hirnschädel von KNM-WT 40000 deutlich kleiner. Wood & Collard (1999) überführten KNM-ER 1470 von Homo in die Gattung Australopithecus. Nach Leakey et al. (2001) sollte er aber eher als Kenyanthropus reklassifiziert werden. (Beide abgebildeten Schädel sind Nachbildungen.) |
|
Die Größe der Kronen der postcaninen Zähne im Verhältnis zum Körpergewicht ist bei „Homo“ habilis ähnlich der von Australopithecus afarensis und Australopithecus africanus; eine Reduktion zu menschlichen Verhältnissen hin ist somit nicht nachweisbar (McHenry & Coffing 2000). Der Kauapparat von „Homo“ habilis und „Homo“ rudolfensis war somit nicht kleiner als der Kauapparat der Australopithecinen. Dagegen besaß Homo erectus einen dem modernen Menschen vergleichbaren Kauapparat.
Ungar et al. (2011) haben fossile Zähne von „Homo“ habilis und Homo erectus miteinander verglichen. Sie stellten fest, dass sich das Zahnabnutzungsmuster der beiden Formen deutlich unterscheidet. Die Forscher schließen daraus, dass sich „Homo“ habilis und Homo erectus unterschiedlich ernährten. Die Ernährung von Homo erectus war vielfältiger, zumindest was die Härte der Nahrung betrifft. Homo erectus ähnelte darin dem heutigen Menschen. Dagegen hat „Homo“ habilis offensichtlich eine weichere Nahrung vergleichbar der von Australopithecus afarensis, Australopithecus africanus und Paranthropus boisei zu sich genommen.
Dauer der Kindheit
Körpermasse, Gehirngröße und das Zahnentwicklungsmuster erlauben Rückschlüsse auf die Kindheitsentwicklung bei Homininen. Danach hatten „Homo“ habilis und „Homo“ rudolfensis eine kurze Kindheit ähnlich wie die Schimpansen (Robson & Wood 2008). Aber auch Homo erectus unterscheidet sich mit einer kürzeren Kindheit vom Entwicklungsmuster des modernen Menschen (Robson & Wood 2008, Dean & Smith 2009).
|
|
Zusammenfassung und Schlussfolgerung
1999 kamen Wood & Collard in einer Übersichtsarbeit zu dem Ergebnis, dass „Homo“ habilis und „Homo“ rudolfensis in wesentlichen Merkmalen dem Anpassungsniveau (Organisationsgrad) der recht menschenaffenartigen Australopithecinen deutlich näher stehen als dem modernen Menschen. Homo erectus ähnelt dagegen in nahezu allen Anpassungsmerkmalen dem modernen Menschen. Wood & Collard gliederten deshalb habilis und rudolfensis aus Homo aus und stuften sie in die Gattung Australopithecus zurück. Diesen Vorschlag hat aber nahezu die gesamte Paläanthropologengemeinschaft ignoriert.
Seit 1999 sind weitere Analysen publiziert worden, die auf die vorgeschlagene Reklassifizierung der beiden Homininen-Arten drängen. „Homo“ habilis zeigt ein Belastungsverhältnis von Vorder- zu Hinterextremitäten ähnlich dem der Schimpansen und ernährte sich wie die Australopithecinen. Bei Homo erectus waren dagegen die Ernährung und das Extremitätenbelastungsmuster menschlich modern.
|
| Dass trotz guter Gründe „Homo“ habilis/rudolfensis nicht umklassifiziert wird, lässt nur eine Vermutung zu: Die morphologische Lücke zwischen den Australopithecinen und Homo überschreitet dann die tolerierbare Grenze. |
|
|
Das Hauptfossil des „Homo“ rudolfensis, der berühmte Schädel KNM-ER 1470, erfuhr eine notwendige Neurekonstruktion mit dem Ergebnis eines etwas kleineren Schädelvolumens und eines stärker vortretenden Gesichtes. Damit rückt „Homo“ rudolfensis noch näher als zuvor an die robusten Australopithecinen heran. Wegen verblüffender Ähnlichkeiten mit dem neu entdeckten Homininen Kenyanthropus platyops wurde vorgeschlagen, KNM-ER 1470 dieser neuen Gattung zuzuordnen. Es gibt somit gute Gründe „Homo“ habilis in Australopithecus habilis und „Homo“ rudolfensis in Kenyanthropus rudolfensis umzuklassifizieren. Dass trotzdem an der alten Klassifizierung festgehalten wird, lässt nur eine Vermutung zu: Die morphologische Lücke zwischen den Australopithecinen und Homo überschreitet dann die tolerierbare Grenze.
„Homo“ habilis/rudolfensis wird seit Jahrzehnten als phylogenetisch vermittelnde Fossilgruppe zwischen den Australopithecinen und dem frühesten echten Menschen (Homo erectus) verstanden. Mit ihrer Rückstufung erscheint die Lücke zwischen den recht großaffenähnlichen Australopithecinen und dem echten Menschen (Homo erectus und spätere Homo-Formen) größer. Auch für den Laien erkennbar ergibt sich damit eine Unterteilung der fossilen Homininen in zwei Gruppen: Menschenähnliche Formen, zu denen Homo erectus und die späteren Homo-Arten gehören, die in der Schöpfungsforschung dem Grundtyp Mensch zugeordnet werden, und die mehr großaffen-ähnlichen Formen Sahelanthropus, Orrorin, Ardipithecus, Kenyanthropus, Australopithecus und Paranthropus, die eine separate Gruppe ohne historisch-verwandtschaftliche Beziehung zum Menschen bilden.
|
Literatur
- Baab KL (2008)
- A re-evaluation of the taxonomic affinities of the early Homo cranium KNM-ER 42700. J. Hum. Evol. 55, 741-746.
- Brandt M (1995)
- Der Ursprung des aufrechten Ganges. Neuhausen-Stuttgart.
- Bromage TG, Kullmer O, Schrenk F, Rosenberger A, Thackeray JF & Hogg R (2007)
- Craniofacial architectural constraints resolve major quandry of human evolution. Conference Poster 3001, Congress of International Association for Dental Research (IADR), 21.-24. 3. 2007, New Orleans.
- Bromage TG, McMahon JM, Thackeray JF, Kullmer O, Hogg R, Rosenberger AL, Schrenk F & Enlow DH (2008)
- Craniofacial architectural constraints and their importance for reconstructing the early Homo skull KNM-ER 1470. J. Clin. Pediatric Dentistry 33, 43-54.
- Carlson K, Wrangham R, Muller M, Sumner R, Morbeck ME, Nishida T, Yamanaka A & Boesch C (2010)
- The locomotor repertoire of early Homo: insights from chimpanzee variation. Am. J. Phys. Anthropol. Suppl. 50, 76.
- Dean MC & Smith BH (2009)
- Growth and development of the Nariokotome Youth, KNM-WT 15000. In: Grine FE, Fleagle JG & Leakey RE (eds) The first humans – origin and early evolution of the genus Homo. Springer, pp 101-120.
- Gebo DL & Schwartz GT (2006)
- Foot bones from Omo: implications for hominid evolution. Am. J. Phys. Anthropol. 129, 499-511.
- Haeusler M & McHenry HM (2004)
- Body proportions of Homo habilis reviewed. J. Hum. Evol. 46, 433-465.
- Haeusler M & McHenry HM (2007)
- Evolutionary reversals of limb proportions in early hominids? Evidence from KNM-ER 3735 (Homo habilis). J. Hum. Evol. 53, 383-405.
- Haeusler M, Schiess R & Boeni T (2011)
- New vertebral and rib material point to modern bauplan of the Nariokotome Homo erectus skeleton. J. Hum. Evol. 61, 575-582.
- Hartwig-Scherer S (1999)
- „Homo“ habilis ab jetzt kein Mensch mehr. Stud. Int. J. 6, 85-87.
- Hartwig-Scherer S & Brandt M (2007)
- KNM-ER 1470 kein Vorfahr des Menschen – Aufstieg und Fall eines Stars. Stud. Int. J. 14, 74-76.
- Hartwig-Scherer S & Martin RD (1991)
- Was “Lucy” more human than her “child”? J. Hum. Evol. 21, 439-450.
- Johanson DC, Masao FT, Eck GG, White TD, Walter RC, Kimbel WH, Asfaw B, Manega P, Ndessokia P & Suwa G (1987)
- New partial skeleton of Homo habilis from Olduvai Gorge, Tanzania. Nature 327, 205-209.
- Jungers WL (2009)
- Interlimb proportions in humans and fossil hominins: variability and scaling. In: Grine FE, Fleagle JG & Leakey RE (eds) The first humans – origin and early evolution of the genus Homo. Springer, pp 93-98.
- Kidd R (1999)
- Evolution of the rearfoot. J. Am. Podiatr. Med. Ass. 89, 2-17.
- Korey KA (1990)
- Deconstructing reconstruction: the OH 62 humerofemoral index. Am. J. Phys. Anthropol. 83, 25-33.
- Leakey REF (1972)
- Evidence for an advanced Plio-Pleistocene hominid from East Rudolf, Kenya. Nature 242, 447-450.
- Leakey MG, Spoor F, Brown FH, Gathogo PN, Kiarie C, Leakey LN & McDougall I (2001)
- New hominin genus from eastern Africa shows diverse middle Pliocene lineages. Nature 410, 433-440.
- Leakey LSB, Tobias PV & Napier JR (1964)
- A new species of the genus Homo from Olduvai Gorge. Nature 202, 7-9.
- Leakey RE, Walker A, Ward CV & Grausz HM (1989)
- A partial skeleton of a gracile hominid from the Upper Burgi Member of the Koobi Fora Formation, East Lake Turkana, Kenya. In: Giacobini G (ed) Hominidae. Proceedings of the 2nd International congress of human paleontology. Milan, pp 167-173.
- McHenry HM & Coffing K (2000)
- Australopithecus to Homo: transformations in body and mind. Annu. Rev. Anthropol. 29, 125-146.
- Moyà-Solà S, Köhler M, Alba DM & Almécija S (2008)
- Taxonomic attribution of the Olduvai Hominid 7 manual remains and the functional interpretation of hand morphology in robust australopithecines. Folia Primatol. 79, 215-250.
- Reno PL, DeGusta D, Serrat MA, Meindl RS, White TD, Eckhardt RB, Kuperavage AJ, Galik K & Lovejoy CO (2005)
- Plio-Pleistocene hominid limb proportions: evolutionary reversals of estimation errors? Curr. Anthropol. 46, 575-588.
- Richmond BG, Aiello LC & Wood BA (2002)
- Early hominin limb proportions. J. Hum. Evol. 43, 529-548.
- Robson SL & Wood B (2008)
- Hominin life history: reconstruction and evolution. J. Anat. 212, 394-425.
- Ruff C (2009)
- Relative limb strength and locomotion in Homo habilis. Am. J. Phys. Anthropol. 138, 90-100.
- Spoor F, Leakey MG, Gathogo PN, Brown FH, Antón S, McDougall I, Kiari C, Manthi FK & Leakey LN (2007)
- Implications of new early Homo fossils from Ileret, east of Lake Turkana, Kenya. Nature 448, 688-691.
- Spoor F, Wood BA & Zonneveld F (1994)
- Implications of early hominid labyrinthine morphology for evolution of human bipedal locomotion. Nature 369, 645-648.
- Susman RL (2008)
- Brief communication: evidence bearing on the status of Homo habilis at Olduvai Gorge. Am. J. Phys. Anthropol. 137, 356-361.
- Trocheri MW, Marzke MW, Liu D, Bae M, Jones GP, Williams RC & Razdan A (2003)
- Functional capabilities of modern and fossil hominid hands: three-dimensional analysis of trapezia. Am. J. Phys. Anthropol. 122, 101-112.
- Ungar PS, Krueger KL, Blumenschine RJ, Njau J & Scott RS (2011)
- Dental microwear texture analysis of hominins recovered by the Olduvai Landscape Paleoanthropology Project, 1995-2007. J. Hum. Evol., doi:10.1016/j.jhevol.2011.04.006.
- Walker A (1976)
- Remains attributable to Australopithecus in the East Rudolf succession. In: Coppens Y, Howell FC, Isaac GL & Leakey REF (eds) Earliest man and environments in the Lake Rudolf Basin. Chicago, pp 484-489.
- Wood BA (2009)
- Where does the genus Homo begin, and how would we know? In: Grine FE, Fleagle JG & Leakey RE (eds) The first humans – origin and early evolution of the genus Homo. Springer, pp 17-28.
- Wood B & Aiello LC (1998)
- Taxonomic and functional implications of mandibular scaling in early hominins. Am. J. Phys. Anthropol. 105, 523-538.
- Wood B & J Baker (2011)
- Evolution in the genus Homo. Ann. Rev. Ecol. Evol. Syst. 42, 47-69.
- Wood B & Collard M (1999)
- The human genus. Science 284, 65-71.
- Wood B & Constantino P (2007)
- Paranthropus boisei: fifty years of evidence and analysis. Yearb. Phys. Anthropol. 50, 106-132.
-
|
|  |
