 |
Über den Ursprung des Insektenflügels
von Reinhard Junker
Studium Integrale Journal
17. Jahrgang / Heft 2 - November 2010
Seite 96 - 99
|
 |
Zusammenfassung: Eine japanische Forschergruppe berichtete kürzlich über entwicklungsgenetische Untersuchungen zur ontogenetischen* Bildung der Insektenflügel (Niwa et al. 2010). Aus ihren Ergebnissen schließen sie, dass einige Bauplanänderungen, die zur Ausbildung des häutigen Insektenflügels erforderlich waren, durch relativ wenige Änderungsschritte möglich gewesen sein könnten. Die von den Autoren vorgestellten Befunde aus der Entwicklungsbiologie sind jedoch keineswegs ausreichend für die Erklärung des evolutionären Werdens von Insektenflügeln, sondern nur Randbedingungen, die beim Versuch einer evolutionstheoretischen Erklärung berücksichtigt werden müssen.
| |
|
Einleitung
 |
| Abb. 1: Apis mellifera, die Honigbiene. Wie entstanden die häutigen Flügel? Foto: Winfried Borlinghaus |
|
Wie entstanden ganz neue Bauelemente der Lebewesen wie z. B. Flügel? Woher kommen evolutive Neuheiten? In den letzten Jahren wurden zahlreiche erstaunliche Entdeckungen in der Entwicklungsbiologie gemacht, von denen man sich Antworten auf diese Frage erhofft. In der Entwicklungsbiologie geht es um Vorgänge, die zum Wachstum und zur Differenzierung des individuellen Organismus führen, ihr Forschungsgegenstand ist also die Entwicklung von der Befruchtung bis zum ausgewachsenen Organismus (Ontogenese). Im Zeitalter der Molekulargenetik geht es dabei besonders um die Rolle der (Regulations-)Gene im Zellwachstum und in der Spezialisierung und Differenzierung der Zellen, in der Ausbildung der Organanlagen und ihrer Ausformung. In der Evolutionären Entwicklungsbiologie (kurz: Evo-Devo) versucht man, aus der Erforschung der Regulationsprozesse in der Ontogenese Hinweise für die hypothetische Phylogenese (Stammesgeschichte, Evolution) zu gewinnen. Zur Evolutionären Entwicklungsbiologie siehe Junker (2009).
Weshalb wird in die evolutionäre Entwicklungsbiologie die Hoffnung gesetzt, dass mit ihrer Hilfe Fragen nach dem Ursprung evolutiver Neuheiten geklärt werden könnten? Es sind unter anderem die Entdeckung von Regulationsgenen, von Mehrfachnutzung von Genen oder ganzen Entwicklungsmodulen sowie des teilweise modularen Aufbaus der Netzwerke zwischen den Erbinformationen und zahlreichen zellulären Prozessen und Entwicklungsvorgängen. Manche Evolutionsbiologen sehen darin das Potential, dass durch Neuverschaltungen von Genen, insbesondere Regulationsgenen, oder ganzer Entwicklungsmodule neue Organe relativ schnell entstehen können. Diese Möglichkeit wird zum Beispiel bei der Entstehung des Schildkrötenpanzers (vgl. den Artikel über die Schildkröten in dieser Ausgabe) oder des Fledermausflügels diskutiert; und jüngst am Beispiel der Entstehung des Insektenflügels, um das es im Folgenden gehen soll.
|
|
Der Ursprung des Insektenflügels
Eine japanische Forschergruppe um Shigeo Hayashi berichtete kürzlich über entwicklungsgenetische Untersuchungen zur ontogenetischen Bildung der Insektenflügel (Niwa et al. 2010). Aus ihren Ergebnissen schließen sie, dass einige Bauplanänderungen, die zur Ausbildung des häutigen Insektenflügels (Abb. 1) erforderlich waren, durch relativ wenige Änderungsschritte möglich gewesen sein könnten. Es handelt sich dabei um ein typisches Beispiel dafür, wie man versucht, die Ergebnisse der Entwicklungsbiologie für die Lösung evolutionstheoretischer Fragestellungen zu nutzen. Es soll in diesem Artikel jedoch gezeigt werden, dass die Erklärungskraft bezüglich evolutionärer Hypothesen sehr begrenzt ist: Die von den Autoren vorgestellten Befunde aus der Entwicklungsbiologie sind keineswegs ausreichend für die Erklärung des evolutionären Werdens von Insektenflügeln, sondern sind nur Randbedingungen, die beim Versuch einer evolutionstheoretischen Erklärung berücksichtigt werden müssen.
Die Entstehung des Insektenflügels konnte durch Fossilfunde nicht erhellt werden, „da die ältesten bekannten Fossilien geflügelter Insekten (Pterygota) bereits über einen hochentwickelten, voll ausgebildeten Flugapparat mit großen Flügeln und komplexer Flügeladerung verfügen“ (Hörnschemeyer 2009). Fossilien, die Übergänge zeigen, sind nicht bekannt. Hypothesen zur evolutiven Entstehung des Insektenflügels beruhen daher auf vergleichenden Studien heute lebender oder fossiler Formen. Zur Entstehung des Insektenflügels werden zwei Hypothesen diskutiert (vgl. Hörnschemeyer 2009, Niwa et al. 2010):
1. Epicoxal-Theorie: Die Flügel sind aus dem beweglichen äußeren Anhang des obersten Beingliedes (der Epicoxa) der ursprünglichen Gliederfüßer-Beine hervorgegangen. Der Vorteil dieser Theorie besteht darin, dass bereits Muskulatur vorhanden ist. Problematisch sind dagegen u. a. die Entstehung von flachen Tragflächen und der rückenseitigen Lage der Flügel.
2. Paranotaltheorie. Die Insektenflügel sind als seitliche Abfaltungen im Brustsegment bzw. im Grenzbereich Rücken-Seite entstanden. Diese Theorie hat den Vorteil, dass die Flächigkeit leichter verständlich ist; ein Problem von vielen ist aber die Entstehung der Flugmuskulatur und der Gelenke, die nach dieser Theorie de novo erfolgt sein müssten.
|
Bei allen Überlegungen zur Entstehung der Flügel muss man sich vor Augen halten, dass Flügel von Insekten immer mehr als relativ simple Ausstülpungen sind, auch bei Insekten in Bernstein und bei den als „ursprünglich“ geltenden Eintagsfliegen. Dabei ist nichts „primitiv“! Besonders erstaunlich ist z. B. die Libellenflügel-Konstruktion. Hier werden die Flügel durch eine äußerst komplizierte Muskulatur unabhängig voneinander bewegt, während bei Wespen, Bienen, Fliegen der Auf- und Abschlag der Flügel synchron durch sich zusammenziehende Thoraxmuskulatur erfolgt. Jede Flügel-Muskulatur-Gelenk-Kombination ist für sich genommen ein technisch fein abgestimmtes Organ. Bei Bienen haken etwa die Hamuli (Häkchen am Vorderrand des Hinterflügels, Abb. 3) von unten (!) in die entsprechend ausgebildete Randleiste des Vorderflügels ein, so dass beim Abschlag mit den größeren Kraftwirkungen der Hinterflügel vom Vorderflügel quasi flächig mit nach unten gedrückt wird. Beim Aufschlag wird der Hinterflügel mit weniger Kraft nach oben gezogen.
Die Konstruktion ist so ausgefeilt, dass wegen der im äußeren Bereich der Flügel auftretenden größeren Kräfte zweckmäßigerweise auch eine drastische Vergrößerung der Auflagefläche des Vorderflügels auf den Hinterflügel vorzufinden ist. Dies wird durch eine Verbreiterung des Vorderflügels im Außenbereich von dessen Hinterkante erreicht. So wird beim Abschlag verhindert, dass sich die Hamuli wie bei einem Reißverschluss vom Vorderflügel lösen. Dazu kommt beim Bienenflügel die speziell gefertigte Muskulatur, die – gesteuert durch das Gehirn – dafür sorgt, dass sich der Hinterflügel nach dem Landen aus der Leiste des Vorderflügels löst und als Ganzes unter den Vorderflügel geschoben werden kann.
|
|
|
Die Untersuchungen von Niwa et al. und ihr Modell
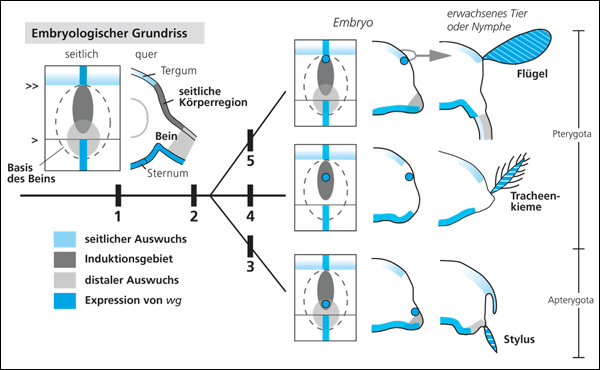 |
| Abb. 2: Embryologischer Grundriss der Segmente der Insekten und ein Modell zur Evolution des Flügels. Links: Seitliche Sicht und Querschnitt eines generalisierten Segments, Expression von wg und Unterteilung in drei Regionen. Rechts: Vorgeschlagenes Szenario der Evolution des Insektenflügels. (1) Etablierung des Segment-Grundplans mit wg-freier seitlicher Region, (2) Erwerb des wg-vg-Moduls zur Induktion der rückenseitigen Abzweigung des Auswuchses, (3) Bildung des Stylus, (4) Bildung der Tracheenkiemen, (5) Bildung des Flügels. Die Position der induktiven wg-vg-Expression variierte in der seitlichen Region (evolutionäre Drift). Eine erfolgreiche Kombination beider Module soll die schnelle Entstehung eines flächigen Flügel-Auswuchses ermöglicht haben. Näheres im Text. Nach Niwa et al. (2010) |
|
Niwa et al. (2010) untersuchten drei für die Flügelausformung wichtige Regulatorgene (wingless: wg, vestigial: vg und apterous: ap) bei Vertretern zweier Insektengruppen, die als evolutionär ursprünglich eingestuft werden, nämlich bei den Eintagsfliegen (Ephemeroptera) und bei ungeflügelten Felsenspringern (Archaeognatha). Diese Insekten besitzen rückenseitig Körperanhänge, und zwar Tracheenkiemen (Nymphen der Eintagsfliege) bzw. sogenannte Styli (Felsenspringer), das sind kurze, stabförmige Auswüchse an der Körperseite von deren Nymphen. Die Untersuchungen der Wissenschaftler ergaben, dass zwei Regulationsmodule, die mit wg und vg verknüpft sind und die für die Ausbildung des Flügels benötigt werden, auch in den flügellosen Insekten vorkommen: Das eine ist im Bereich der seitlichen Körperregion aktiv und induziert stabförmige Beinanhänge, das andere im Bereich der Grenze zwischen Rücken und Seite und ist mit dem flachen Auswuchs in deren Grenzbereich verbunden.
Niwa et al. (2010, 174) schlagen folgendes Modell der Flügelentstehung vor: Zwei Regulationsgene, die in der ontogenetischen Entwicklung der Flügel beteiligt sind (nämlich wg und vg), hatten in den mutmaßlichen Vorfahren der geflügelten Insekten eine andere Funktion, nämlich in der Induktion des oberen stäbchenförmigen Asts der verzweigten Gliedmaßen bzw. in der Induktion von flachen Auswüchsen an der Grenze von Rückenschild und seitlicher Körperregion (Abb. 2). Durch Kombination dieser beiden Entwicklungsmodule und die Verschiebung ihres Wirkungsbereichs in der seitlichen Körperregion sei die rasche Entstehung des Flügels begünstigt worden. Dadurch sei außerdem die flache Form der seitlichen Körperauswüchse mit der Beweglichkeit der Gliedmaßen kombiniert worden.
Dieses Modell ist zugleich eine Kombination der Epicoxal- und der Paranotaltheorie (s. o.). Typisch für „Evo-Devo“ ist die Idee, dass sich durch wenig geänderte Verschaltungen von Genen und ihrer Wirkungsbereiche neue Organe bilden könnten (hier der Flügel von Insekten) und darüber hinaus, dass dies in relativ kurzen Zeitabschnitten vonstatten gehen könnte (womit sich die Suche nach Übergangsformen erübrigen würde). So spricht Hemminger (2010) von einer „einfachen Integration“ der beiden vorher getrennten Steuer-Module und kommentiert: „Das vorliegende Modell zeigt, dass sich selbst große Bauplanänderungen, … auf der Ebene der Entwicklungssteuerung durch relativ wenige Veränderungsschritte erklären lassen“ (Hervorhebung nicht im Original).
|
|
|
Was haben Niwa et al. gezeigt?
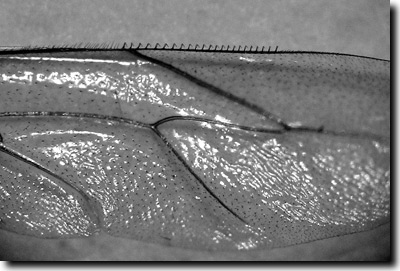 |
| Abb. 3: Hamuli des linken Hinterflügels der Honigbiene Apis mellifera (von ventral) – eine ausgeklügelte Arretierungsvorrichtung. Foto: Winfried Borlinghaus |
|
Bei der hier anstehenden Frage, wie ein Organ entsteht, geht es um Mechanismen und Abläufe. Die Untersuchungen von Niwa et al. haben aufgedeckt, dass für die Induktion des Stylus bei Felsenspringern, der Tracheenkiemen bei Eintagsfliegenlarven und bei den Flügeln der Taufliege teilweise gleiche Entwicklungsmodule genutzt werden – zweifellos eine sehr interessante Entdeckung durch wegweisende Experimente. Wie diese gleichen Module für die verschiedenen Aufgaben – hier die Induktion verschiedener Körperanhänge – rekrutiert und verschaltet wurden, wird durch diese Gemeinsamkeiten aber überhaupt nicht geklärt. Die Befunde von Niwa et al. liefern also eine wichtige Basis für das von ihnen vorgeschlagene Entstehungsmodell. Das Verständnis der ontogenetischen Zusammenhänge ist aber als solches nicht schon die Erklärung des hypothetisch vorgeschlagenen Evolutionsvorgangs. Die Autoren haben Bausteine geliefert, die berücksichtigt werden müssen, wenn man konkrete Hypothesen über evolutive Mechanismen des Umbaus hin zum Insektenflügel aufstellt. Sie äußern sich dabei selbst vorsichtig und unterscheiden zwischen den gewonnenen Daten und den Spekulationen über mögliche Evolutionswege.
Eine Erklärung müsste zeigen, wie durch ungerichtete Mutationen die postulierten Veränderungen eintreten, es bräuchte experimentelle Belege dafür, dass solche Verschaltungen von Modulen (auch schrittweise) möglich, funktional und selektionspositiv sind (andernfalls könnten sie nicht durchsetzen). Dabei muss natürlich nicht gleich die Flugfunktion erreicht werden, aber hypothetische Zwischenstufen auf dem Weg zur Flugfähigkeit müssen irgendeine Funktion ausüben. Weiter müsste geklärt werden, welche Nebenwirkungen die vermuteten Neuverschaltungen hatten und ob sie tolerabel sind. Dieselben Gene (insbesondere die Masterkontrollgene) üben örtlich und zeitlich koordiniert zahlreiche Funktionen im sich entwickelnden Organismus aus und sind in einem komplizierten Netzwerk miteinander verbunden. Änderungen betreffen also viele Aspekte der Organisation der Lebewesen und ihrer Ontogenese.
 |
Die postulierte Neuverknüpfung
von Entwicklungsmodulen benötigt
vielfache parallele Abstimmungen. |
 |
|
Kurz: die postulierte Neuverknüpfung von Entwicklungsmodulen benötigt vielfache parallele Abstimmungen, damit sie selektionspositiv bzw. mindestens neutral sein kann. Mit „einfachen“ Änderungen auf der Ebene der Entwicklungssteuerung ist es deshalb bei weitem nicht getan.
Die Integration eines neuen bzw. die Abwandlung eines vorhandenen Schalters in einem örtlich und zeitlich fein abgestimmten Netzwerk von Entwicklungsmodulen erzeugt nicht per se – wie übrigens auch in der Technik nicht – eine neue Funktionalität oder ein neues Bauprinzip des betroffenen Teilsystems.
 |
| Abb. 4: Relativ frisch geschlüpftes Weibchen der Becher-Azurjungfer (Enallagma cyathigerum). Ein einziges Libellenflügelgelenk mit der entsprechenden Muskulatur versehen ist dermaßen komplex aufgebaut, dass umfangreiche wissenschaftliche Arbeit erforderlich ist, um allein die Freiheitsgrade des möglichen Flügel- Bewegungsablaufs technisch zu verstehen, die Kraftübertragung mathematisch zu beschreiben und im Modell umzusetzen. Foto: Winfried Borlinghaus |
|
Doch selbst wenn die mit der Verflechtung der Gene und Entwicklungsmodule verbundenen Probleme der Steuerung gelöst werden könnten, hätte man mit der Rekrutierung und Verschaltung zweier Regulationsgene und der Verschiebung ihres Wirkungsbereichs nicht einmal ansatzweise die Entstehung des Flügels erklärt. Denn der Flügel muss eine bestimmte Form besitzen, die Flügelmuskulatur muss entsprechend angepasst sein, die Verbindung des Flügels mit der Flugmuskulatur muss funktional sein, Gelenke müssen ausgebildet sein, das Insekt benötigt ein entsprechendes Verhalten und alle diese Vorgänge benötigen neue Steuerungsvorgänge in der Ontogenese. Ein Zitat zum Aufbau des Flügelgelenks kann die Problematik beispielhaft verdeutlichen: „Es handelt sich dabei um ein hoch komplexes System aus mehreren Skleriten, die in ihrem mechanischen Zusammenspiel dafür sorgen, dass die Bewegungen der Muskulatur auf den Flügel übertragen werden. Dazu gehören neben dem Auf- und Abschlag auch Steuerbewegungen und das Zusammenlegen oder Falten der Flügel“ (Hörnschemeyer 2009). Neben den bereits genannten organismusinternen Rahmenbedingungen eines Systemumbaus („constraints“) bleibt im theoretischen System der Synthetischen Evolutionstheorie die Frage offen, welche Selektionsdrücke den postulierten Umbau steuerten. Wurde in Richtung auf die Flugfähigkeit hin selektiert, wären die o. g. Eigenschaften des Flügels relativ kurzfristig und gleichzeitig erforderlich gewesen, um selektionspositiv zu sein. Wurde nicht in Richtung Flugfähigkeit selektiert, bleibt zu klären, welche selektionspositive Primärfunktionen die „Vor“-flügel besaßen, bevor diese zum Fliegen genutzt wurden und wann, warum und wie der Funktionswechsel erfolgte.
Ein einfacher und schneller Erwerb der Flugfähigkeit ist durch die Befunde von Niwa et al. also nicht einmal ansatzweise demonstriert. Die populäre Darstellung „But it turns out that just two genes may explain insect wings“ von Dan Vergano (2010) in USA Today ist durch die Daten nicht im Entferntesten gedeckt. Die Autoren des Originalartikels sind im Gegensatz zur Darstellung bei Hemminger (2010) so vorsichtig, dass sie nur davon sprechen, dass die hypothetische Kombination der beteiligten Regulationsgene eine schnelle Entstehung des Flügels „erleichtert“ habe (S. 174). Macht man sich klar, was man für einen funktionsfähigen Flügel im Einzelnen benötigt, wird deutlich, dass diese „Erleichterung“ gegenüber den noch zusätzlich erforderlichen Änderungen und Innovationen auf dem Weg zum Insektenflügel nur als eine minimale Rahmenbedingung und nicht als Lösung des evolutionstheoretischen Problems betrachtet werden kann.
Dank: Wertvolle Hinweise erhielt ich von Winfried Borlinghaus, Jean-Luc Murk und Henrik Ullrich.
|
|  |
