 |
Einschränkung durch Evolution?
von Niko Winkler
Studium Integrale Journal
11. Jahrgang / Heft 2 - Oktober 2004
Seite 80 - 82
|
|
Unumkehrbare Evolution
„Ein weithin akzeptiertes evolutives Prinzip ist“, so schreiben Zufall und Kollegen (2004), „daß adaptive Anpassung die Möglichkeiten zukünftiger evolutiver Anpassungen einschränkt.“ Eine Reihe von Beobachtungen führte bereits 1890 zur Formulierung eines „Gesetzes“, benannt nach dem Paläontologen Luis Dollo. Dieses Gesetz besagt, daß Evolution nicht umkehrbar ist. Interessanterweise ergibt die relevante Literatur, daß mehr oder weniger ausnahmslos der Verlust von Merkmalen diskutiert wird. Dieser an sich schon interessante Sachverhalt wird nun von Zufall und Kollegen (2004) durch ein Beispiel anhand der Blütenfarben bei Windengewächsen konkretisiert. Die Überschrift des Fachartikels in der Zeitschrift Nature lautet dementsprechend: „Genetische Veränderungen in Verbindung mit Blütenanpassungen schränken das zukünftige evolutive Potential ein.“
Worum geht es? Winden der Gattung Ipomoea haben ursprünglich eine blauviolette Blütenfarbe. Eine kleine Gruppe offensichtlich eng verwandter Gattungen besitzt nun stattdessen rote Blüten, die nun von Kolibris besucht werden. Vögel „fliegen“ auf rote Blüten, während die blauvioletten Arten vorwiegend von Bienen und Hummeln aufgesucht werden. Die Änderung der Blütenfarbe ist nicht der einzige Unterschied zugunsten eines anderen Bestäubers, auch die Blütenform (längere Röhre), der Nektar (zunehmend flüssiger) und andere Merkmale haben sich verändert.
|
|
Farbstoffsynthese in Winden-Blüten
Für das Verständnis der erforschten Vorgänge ist es nötig, sich ein wenig mit der Synthese von Blütenfarbstoffen in diesen Pflanzen zu beschäftigen: Die Blütenfarbe wird bei Ipomoea-Arten zum größten Teil von der An- bzw. Abwesenheit bestimmter an Zucker gekoppelter Blütenfarbstoffe, den Anthocyanen, verursacht. So bewirkt der Blütenfarbstoff Cyanidin-Glucosid blauviolette und der Farbstoff Pelargonidin-Glucosid rote Farben. Der chemische Unterschied zwischen beiden Blütenfarben ist an sich gering, die blauviolette Variante besitzt lediglich eine bestimmte chemische Gruppe (Hydroxylgruppe; –OH) mehr als der rote Farbstoff. Die Anlagerung dieser chemischen Gruppe stellt einen Verzweigungspunkt in der Farbstoff-Synthese dar. Das heißt ab diesem Zeitpunkt unterscheiden sich alle nachfolgenden Farbstoffvorstufen durch jeweils das Vorhandensein der OH-Gruppe (später blauviolett) bzw. der Abwesenheit jener Gruppe (führt zu rot). Durch entsprechende Experimente über künstliche Mutanten konnte gezeigt werden, daß Enzyme von Pflanzen mit blauen Blüten in der Lage sind, auch die rote Variante des Farbstoffs zu erkennen und zu verarbeiten. Die betreffenden Stoffwechsel-Enzyme sind also Generalisten und unterscheiden nicht zwischen rot oder blau. Soweit, so blau.
|
|
Verlust & Degeneration
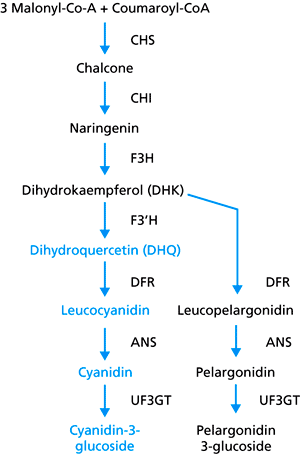 |
| Abb. 2: Der zentrale Teil des Biosynthesewegs des Anthocyanins. CHS Chalcone-Synthase; CHI Chalcon-Isomerase; F3H Flavonon-3-Hydroxylase. Die Zwischenprodukte und Produkte des Cyanidin-Zweiges sind blau gesetzt, die schwarzen (rechts) gehören zum Pelargonidin-Zweig. (Nach Zufall & Rausher 2004) |
|
Um nun zu rotblühenden Pflanzen zu gelangen, würde es genügen, an die Vorstufen des Farbstoffs keine Hydroxylgruppe mehr anzulagern und somit nur die rote Variante zu produzieren. Tatsächlich ist das auch der Fall. In rotblühenden Arten ist nun – erstens – genau jenes Hydroxyl-anlagernde Enzym nicht mehr aktiv. Die Deaktivierung beruht, Versuchen zufolge, nicht direkt auf einem Verlust der enzymatischen Aktivität (wie man im Reagenzglas feststellen kann). Aus unbekannten Gründen funktioniert das Enzym aber in der Pflanze nicht mehr. Des weiteren – zweitens – ist die Ableserate des Gens für dieses Enzym drastisch reduziert.
Überraschend kam nun der dritte Befund, daß die nachfolgenden Enzyme in der Kette der Farbstoff-Produktion durch weitere Veränderungen (Mutationen) nicht mehr in der Lage sind, die Vorstufen für das blaue Pigment zu verarbeiten, selbst wenn diese vorhanden wären. (Wir erinnern uns, in blauvioletten Blüten können die Enzyme Vorstufen für blaue oder rote Farben gleichermaßen verarbeiten.) Um in einem Beispiel zu sprechen wäre es so, als ob ich eine Taschenlampe funktionsunfähig mache, indem ich die Birne herausschraube, die Batterien entnehme und den Schalter demontiere. Ein Ereignis hätte dabei völlig genügt. Gleichermaßen läßt sich in den Blüten feststellen, daß das Hydroxyl-anlagernde Enzym funktionsunfähig geworden ist, dann aber sowieso nicht mehr hergestellt wird, und nachfolgende Enzyme auch nur die Vorstufen für rote Farbe verarbeiten können. Die Autoren der Studie schließen daraus, daß nachdem ein Ereignis zur Änderung von Blauviolett nach Rot eingetreten war, der Stoffwechselweg für Blauviolett durch Mutationen weiter degenerierte. Sie schreiben: „Diese Degeneration macht es sehr unwahrscheinlich, daß I. quamoclit [eine rotblühende Art] je fähig sein wird, Cyanidin-basierte blauviolette Farbpigmente wieder (zurück) zu evolvieren, denn das würde mindestens zwei, eher drei oder mehr [...] gleichzeitige Mutationen benötigen. Das Potential für eine evolutive Veränderung dieses Merkmals scheint also in I. quamoclit als Folge einer Anpassung durch die genetischen Änderungen eingeengt zu sein“ (Zufall et al. 2004).
|
|
Evolutiver Aufbau neuer Funktionen fraglich
Diese Feststellung erinnert sehr an die von Scherer (1996) postulierten Basisfunktionszustände: Der Aufbau von Funktionen, die mehr als 3 (und unter optimistischen Annahmen höchstens 6) Mutationsereignisse voraussetzen, dürfen als nahezu ausgeschlossen gelten, so seine damalige These. Dagegen sind dem Abbau von Funktionen keine Grenzen gesetzt. Dies geht nahtlos in die allgemein anerkannte Feststellung über, daß Artbildung mit einem globalen Verlust von Variabilität (auf genetischer Ebene) einhergeht. Die Frage nach der Herkunft des Stoffwechselweges für blauviolette Blüten im Laufe einer allgemeinen Evolution wird auch in unserem betrachteten Beispiel von den Autoren nicht gestellt. Eine Antwort darauf ist auch nicht bekannt.
 |
„Wir verstehen es nicht,
Neuigkeiten aus dem Nichts
zu schaffen – das ist eine Frage
für Biologen in der Zukunft.“ |
 |
|
Ungewöhnlich ehrlich heißt es zu einem ähnlichen Thema (Entstehung des ersten Lebens), wiederum in der Zeitschrift Nature: „Die Idee einer Gemeinschaft [von Lebewesen] mit gemeinsamer hochentwickelter Biochemie wirft eine weitere Frage auf: Wie evolvierte all dies? ‘Das ist von jemandem anders zu beantworten’, sagt Woese, ‘Wir verstehen es nicht, Neuigkeiten aus dem Nichts zu schaffen – das ist eine Frage für Biologen in der Zukunft’“ (Whitfield 2004). Vielleicht ist diese Frage aber schon längst in einem Klassiker der Weltliteratur beantwortet worden?
|
Literatur
- Zufall RA & Rausher MD (2004)
- Genetic changes associated with floral adaptation restrict future evolutionary potential. Nature 428, 847-850.
- Scherer S (1996)
- Entstehung der Photosynthese. Neuhausen-Stuttgart, 2. erweiterte Auflage.
- Whitfield (2004)
- Borne in a watery commune. Nature 427, 674-676.
-
|
|  |

