 |
|
von Reinhard Junker
Studium Integrale Journal
11. Jahrgang / Heft 2 - Oktober 2004
Seite 59 - 66
|
 |
Zusammenfassung: Das berühmte Ichthyostega aus dem oberen Oberdevon hat seinen Status als „erstes Amphib“ aufgrund neuer Erkenntnisse möglicherweise eingebüßt. Es ist zwar eindeutig ein Tetrapode, lebte aber dennoch wahrscheinlich vornehmlich im Wasser. Der gleichalte Tetrapode Acanthostega mit acht Fingern gilt sogar als ausschließlich wasserlebend. Mit dieser Einschätzung geht ein Paradigmenwechsel in den Evolutionsvorstellungen zur Entstehung der Tetrapoden einher: Die fünfgliedrige Vierbeiner-Extremität dürfte demnach bereits im Wasser entstanden sein, nicht erst bei der Eroberung des Landes.
Aus dieser Sicht folgt jedoch ein bislang ungelöstes evolutionstheoretisches Dilemma: Manche Merkmale sprechen deutlich für eine primäre Lebensweise im Wasser (wonach wichtige Tetrapodenmerkmale also im Wasser entstanden wären), ebenso gewichtige Merkmale deuten jedoch darauf hin, daß sie von landlebenden Formen abstammen, also sekundär wasserlebend sind (wobei dann die Herkunft der landlebenden Vorläufer völlig im Dunkeln wäre).
Die evolutionären Zusammenhänge der Formen des Oberdevons untereinander und mit jüngeren Tetrapoden des Karbons sind zudem unklar.
| |
|
Ichthyostega („Fisch-Panzer“, Abb. 11) ist der bekannteste devonische Tetrapode und als „erstes Landwirbeltier“ in die Geschichte der Paläontologie eingegangen. In der Tat gehört Ichthyostega zu den geologisch frühesten eindeutigen Vierbeinern, da die Hinterextremitäten eindeutig typisch tetrapodenartig ausgebildet sind.
Kopf. Das etwa 1,5 m große Ichthyostega wurde im Famennium (oberes Oberdevon) Grönlands gefunden (Abb. 12). Der Schädel ist ca. 25 cm lang und weist ein ausgeprägtes Mosaik von Fisch- und Tetrapodenmerkmalen auf, besitzt aber auch einige einzigartige Merkmale. Ungewöhnlich ist der im Vergleich mit Fischen und anderen Vierbeinern extrem enge Bau des Hirnschädels. Sein hinterer Teil ist stark verlängert. Die Regionen beidseitig des Hirnschädels scheinen hohle Kammern gewesen zu sein, deren Seiten durch Knochenwände verstärkt waren. Clack konnte im Jahr 2003 durch detaillierte Studien zeigen, daß es sich um ein hochspezialisiertes Ohr handelt, welches wahrscheinlich für ein Hören im Wasser geeignet war. Die Ohrregion einschließlich dem dünnen Stapes (entspricht dem Steigbügel, einem Gehörknöchelchen bei Säugetieren) ist in vielerlei Hinsicht deutlich verschieden von allen bekannten Konstruktionen früher Tetrapoden und tetrapodenähnlichen Fischen wie Eusthenopteron und Panderichthys (Clack 2003; vgl. 1. Teil in der vorigen Ausgabe).
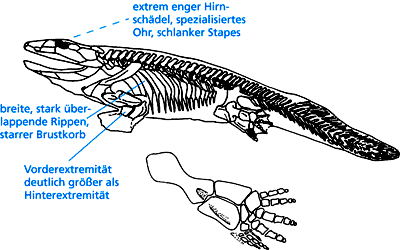 |
| Abb. 11: Ichthyostega; die besonders ausgefallenen Merkmale sind beschriftet. Der Stapes entspricht dem Steigbügel, einem Gehörknöchelchen bei den Säugetieren. (Nach Clack 2002) |
|
Neuerdings wurde die Existenz von Kiemenbögen nachgewiesen (Clack 2003), was ebenfalls (wie die Ohrregion) dafür spricht, daß Ichthyostega mehr ans Wasserleben angepaßt war, als lange Zeit angenommen wurde, auch wenn die Serie der Opercularknochen (Kiemendeckel) größtenteils fehlt (Carroll 1993, 175).
Übriges Skelett. Ein weiteres auffälliges einzigartiges Merkmal, das unter den frühen Tetrapoden sonst völlig unbekannt ist, ist der massive, starre Brustkorb mit überlappenden Rippen (vgl. Abb. 11). Die Rippen sind in der Rumpfregion so stark verbreitert, daß eine Rippe drei oder vier dahinterliegende überlappte, wodurch ein schweres, kaum bewegliches Korsett entstand. Dadurch war eine sinusartige Bewegung nicht möglich. Die Funktion der starren Rippen ist unbekannt und rätselhaft.
Markant ist die typische Tetrapodenextremität, die jedoch nicht 5, sondern 7 Finger aufweist (es sind nur die Finger der Hinterextremität überliefert; s. Abb. 11). Jarvik (1996) bestreitet allerdings die Existenz von 7 Fingern und vertritt die Auffassung, es handle sich nur um 5 echte Finger. Die Hinterextremitäten sind mehr paddel- als beinartig. Der Oberschenkelknochen ist flach, das Sprunggelenk relativ inflexibel und besteht aus weniger Fußwurzelknochen als bei modernen Vierbeinern (Clack 2002, 117). Der Beckengürtel ist groß und robust (Clack 2002, 116).
Die Vorderextremitäten waren kräftig, mit L-förmigem, ziemlich abgeflachtem Oberarmknochen und ziemlich kurzer Speiche und kurzer Elle; damit könnte die Kraft zur Bewegung auf dem Land vermittelt worden sein (Clack 2002, 119). Die Schultern waren massiv, gemessen an der Körpergröße. Der Bau des Schultergelenks und des Ellenbogens dürfte wenig Bewegungsmöglichkeiten erlaubt haben (Clack 2002, 115). Handgelenk und Fingerknochen sind unbekannt.
Die Vorderextremität war viel länger als die Hinterextremität, was für Vierbeiner untypisch ist, da die Verhältnisse sonst genau umgekehrt sind, weil die Hinterextremitäten die Kraft beim Gehen aufbringen müssen.
Der Schwanz ist fischartig gebaut und offenkundig zum Schwimmen da. Er besaß knöcherne Flossenstrahlen, die wie bei Fischen ein Netz unterstützten; bei anderen Tetrapoden ist eine solche Ausprägung bisher unbekannt (Clack 2002, 115).
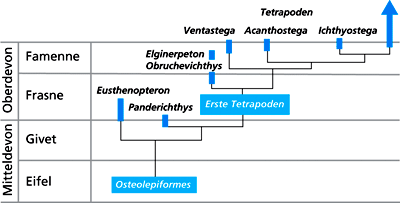 |
| Abb. 12: Stratigraphischer Überblick über die oberdevonischen Tetrapoden. (Nach Clack 2002) |
|
Gesamteindruck und Lebensweise. Läßt man die geschilderten Merkmale Revue passieren, so wird deutlich, daß Ichthyostega vornehmlich im Wasser gelebt haben muß. In diesem Zusammenhang ist auch der Befund wichtig, daß Ichthyostega ausschließlich zusammen mit Fischen gefunden wurde (Clack 2002, 210). Der Status als „erstes Amphib“ ist in den letzten Jahren zunehmend in Frage gestellt worden. Die jüngst erfolgte weitgehende Aufklärung des Baus und der vermutlichen Funktionsweise der Ohrregion stellt der vorläufig letzte Baustein in dieser Entwicklung dar, die – bildhaft gesprochen – Ichthyostega „zunehmend im Wasser versinken läßt“. Das Bild von Ichthyostega hat sich also gegenüber langjährigen Lehrbuchdarstellungen erheblich gewandelt (Clack 2002, 117).
Insgesamt war Ichthyostega in vielfacher Hinsicht einzigartig gebaut und ein „sehr seltsames“ Tier; viele Merkmale finden sich weder bei einem anderen Tetrapoden noch bei einem Fisch (Clack 2002, 115). Allerdings: Wenn auch Ichthyostega weit mehr für ein Leben im Wasser gebaut ist als früher angenommen, sind doch manche Teile des Bauplans für diesen Lebensraum schwer verständlich, so der massive, starre Brustkorb. Außerdem ist unklar, weshalb die Vorderextremitäten deutlich größer sind als die Hinterextremitäten. Dienten sie dazu, sich am Ufer kurzfristig ein Stück weit hochzuziehen?
 |
Der Körperbau von Ichthyostega
entspricht nicht dem, was man von
einem primitiven Tetrapoden erwartet. |
 |
|
Das postkranielle Skelett (Skelett nach dem Schädel) weist sehr beachtliche Ähnlichkeiten mit dem von Seehunden auf; auch Parallelen zum Flußdelphin werden diskutiert (Coates & Clack 1995, 373; Clack 2002, 119); es wird spekuliert, ob die Finger Schwimmhäute besaßen. Bemerkenswerte Ähnlichkeiten von Ichthyostega und Seehunden sind: Brustflosse als Stütze, Hinterbeine als Antriebspaddel; die Oberarm- und Oberschenkel-Proportionen und ein schmaler Beckengürtel (Coates & Clack 1995, 379).
Evolutionäre Betrachtungen. Clack (2003, 65) bezeichnet die Morphologie von Ichthyostega als „rätselhaft“; sie entspreche nicht dem, was man von einem sehr primitiven Tetrapoden erwarte. Die ausgeprägten Spezialisierungen lassen Ichthyostega als blinden Seitenzweig erscheinen.
Als Übergangsform zwischen Fischvorfahren und eindeutigen Landtetrapoden ist Ichthyostega aus folgenden Gründen ungeeignet (vgl. Tab. 1):
 |
| Tab. 1: Einige wichtige Merkmale von Ichthyostega |
|
- Das Ohr von Ichthyostega (inklusive Stapes) ist „vollkommen einzigartig“ und unterscheidet sich markant vom zeitgleichen Acanthostega. Ebenso unterscheidet sich die Ohrregion grundlegend von der Situation bei potentiellen Vorfahren wie anderen frühen Tetrapoden (Clack 2003, 69).
- Die Rippen sind stark überlappend und in dieser Hinsicht ganz einzigartig gebaut.
- Die Vorderextremitäten sind viel größer als die Hinterextremitäten, während die Verhältnisse bei anderen frühen Tetrapoden umgekehrt sind.
- In der postkraniellen Anatomie erscheint Ichthyostega „fortschrittlicher“ als Acanthostega, in der Anatomie des Hirnschädels und der Kiefer ist es dagegen umgekehrt (Clack 2002, 155) – ein ausgeprägter Merkmalswiderspruch.
|
|
Acanthostega
Heute oft mit einem Atemzug mit Ichthyostega wird Acanthostega („Spitz-Panzer“) genannt (Name nach einem markanten Zacken am hinteren Ende des Schädeldachs; vgl. Abb. 13). Gemeinsam ist beiden zwar das geologische Alter, das Fundgebiet (Grönland) und der Besitz einer tetrapodenartigen Extremität, doch in vielerlei Hinsicht unterscheiden sich beide Gattungen deutlich. Von „primitiven“ Ähnlichkeiten abgesehen haben Ichthyostega und Acanthostega fast nichts gemeinsam (Clack 2002, 121).
2/3 der Merkmale von Acanthostega werden zwar als spezielle (abgeleitete) Merkmale von Vierbeinern (Tetrapoden-Autapomorphien) gewertet (Clack & Coates 1995, 359), darunter die typische Tetrapoden-Extremität, dennoch war Acanthostega wasserlebend, wahrscheinlich ausschließlich. Diese Gattung trug wesentlich zur Erkenntnis bei, daß der Besitz einer tetrapodenartigen Extremität nicht mit einer (teilweisen) Lebensweise an Land gleichzusetzen ist. (Wie oben gezeigt wurde, unterstützt Ichthyostega diese Erkenntnis mittlerweile ebenfalls.)
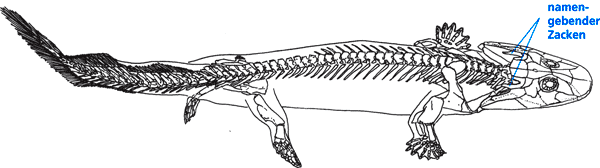 |
| Abb. 13: Acanthostega mit 8 Fingern (Nach Clack 2002) |
|
Kopf. Acanthostega war mit 20 cm Schädellänge kleiner als Ichthyostega. Die Schnauze ist flach und abgerundet. Das Seitenlinienorgan am Kopf (zur Orientierung im Wasser) ist markanter als bei Ichthyostega (Clack 2002, 121). Die Bezahnung gleicht insgesamt zeitgenössischen Fleischflosser-Fischen und keinem Tetrapoden (Clack 2002, 122/4; Clack 1994, 16). Der Schädel ist mit dem Schultergürtel relativ fest verbunden.
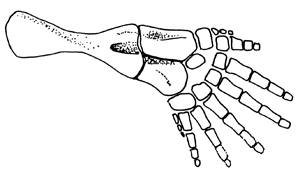 |
| Abb. 14: Hinterextremität von Acanthostega (Nach Clack 2002) |
|
Der Hirnschädel gleicht zwar Fischen wie Eusthenopteron, Panderichthys oder auch frühen Strahlflosser-Fischen (Clack 1994, 393), doch sind gegenüber Panderichthys die Ohrregion und der mittlere Teil des Hirnschädels deutlich anders gebaut (Clack 2001, 393f.). Diese schnellen Änderungen bestimmter Teile des Hirnschädels (vergl 1. Teil über Panderichthys) kontrastieren mit der konservativen Natur anderer Teile (Clack 2001, 395).
Der kräftige Stapes (entspricht dem Steigbügel, s. o.) ist fest mit der Wand des Hirnschädels verbunden und fungiert als einzige Verbindung zwischen Ohrkapsel und Gaumen. Er hat also keine Funktion in der Schallübertragung und unterscheidet sich deutlich vom leichten, schlanken Stapes der meisten frühen Tetrapoden (Clack 1994, 392). Es gleicht eher dem Hyomandibulare, einem Knochen im Schädel-Kiemenbereich bestimmter Quastenflosser (Actinistier) und devonischen Lungenfischen (Janvier 1996, 272). Es hat wohl eine vergleichbare Funktion, nämlich der Kontrolle von Gaumen- und Spritzloch-Bewegung beim Atmen (Clack 1989, 427). Dies paßt nicht zu einer Übergangsstellung von Acanthostega; Übergangsformen fehlen (Janvier 1996, 272).
Der Schultergürtel ist insgesamt fischartiger als bei Ichthyostega (Clack & Coates 1995, 363). Eine für die Kiemenatmung nützliche Serie von Knochen, der Operculogular-Apparat, fehlt zwar, doch dies ist auch bei zahlreichen Fischen der Fall. Acanthostega ähnelt im Kiemenskelett kiemenatmenden Lungenfischen. Beide haben ein Kiemenskelett und einen Schultergürtel mit einer breiten Knochenfläche („postbranchiale Lamina“; wichtig für Kiemenatmung); sehr wahrscheinlich hatte Acanthostega daher innere Kiemen (Coates & Clack 1991, 235).
Übriges Skelett. Die Vorderextremität war vermutlich seitwärts gerichtet, wohl aber beweglicher als bei Ichthyostega (Clack 2002, 124). Der Oberarmknochen ist breit L-förmig – typisch für frühe Tetrapoden –; Speiche und Elle unterscheiden sich jedoch in verschiedener Hinsicht von allen anderen Tetrapoden. Die Speiche ist am körpernahen Ende des Oberarmknochens zylindrisch, körperfern aber abgeflacht und ca. doppelt so lang wie die Elle. Letztere ist flach und hat am körperfernen Ende keine Gelenkflächen; das Handgelenk war wahrscheinlich kaum beweglich (Clack & Coates 1995, 363f.). Insgesamt wirken die Extremitäten eher als Paddel denn als Füße und waren gut zum Schwimmen konstruiert. Die Hinterbeine (Abb. 14) waren etwas größer als die Vorderextremitäten (also umgekehrte Verhältnisse wie bei Ichthyostega).
Auch die Knochen der Hinterbeine waren abgeflacht, die Extremität insgesamt paddelförmig und vermutlich passend fürs Schwimmen konstruiert (Clack 2002, 127). Zum Tragen des Gewichts waren die Extremitäten kaum geeignet; die genauen Bewegungsmöglichkeiten sind allerdings schwer einzuschätzen (Clack 2002, 137).
 |
| Abb. 15: Rekonstruktion von Acanthostega als schwimmender Vierbeiner. (© Richard Hammond) |
|
Die Wirbelsäule ist von vorne bis hinten auffallend gleichförmig, ähnlich wie bei Fischen wie Eusthenopteron. Die Rippen sind viel kürzer und ganz anders gebaut als bei Ichthyostega, was auf eine ganz andere Lebensweise hinweist (Clack 2002, 126). Der Beckengürtel besteht aus einem einzelnen Element und ist relativ klein für einen Tetrapoden, jedoch groß für einen Fisch und in mancher Hinsicht tetrapodenähnlicher als fischähnlich. Eine Verbindung mit der Wirbelsäule bestand vermutlich nur durch Bänder (Clack 2002, 126). Die Hüfte konnte dadurch kaum das Körpergewicht tragen.
Die Schwanzflosse ist lungenfischähnlich gebaut. Sie konnte Muskelwellen durch den ganzen Körper erzeugen (Coates 1996, 404). Flossenstrahlen sind länger und zahlreicher als bei Ichthyostega. Es handelt sich um den Schwanz eines lauernden Räubers; er paßt nicht zu einem Tier, das regelmäßig Landgänge unternimmt.
Lebensweise. Clack (2002, 124) stellt als Gesamteindruck fest: Fast alle Merkmale von Acanthostega legen ein ausschließliches Wasserleben nahe (vgl. Tab. 2; Abb. 15). Abgesehen vom Besitz von Fingern gilt: Viele Merkmale von Acanthostega, die in einen Zusammenhang mit Landleben gebracht werden, kommen auch bei modernen Fischen vor, die keine Beziehungen zu einem Leben an Land haben (Clack 2002, 137). Offenbar ist aber auch der Besitz von Fingern kein untrügliches Indiz für ein (wenigstens teilweises) Landleben.
Vermutlich lebte Acanthostega in vegetationsreichen Uferzonen, in denen eine tetrapodenartige Extremität passend war. Wie alle anderen oberdevonischen Tetrapoden ist auch Acanthostega ausschließlich zusammen mit Fischen fossil überliefert (Clack 2002, 110).
Eine gehende Fortbewegung im Wasser ist übrigens nicht so ungewöhnlich: sie wird auch vom heute lebenden Sargassum-Froschfisch (Histrio) praktiziert (Ahlberg & Milner 1994, 510).
 |
| Tab. 2: Merkmale, die auf ein dauerndes Wasserleben von Acanthostega hinweisen. |
|
Evolutionäre Betrachtungen. Wie bereits erwähnt, trug Acanthostega maßgeblich zu einem Umdenken bezüglich der Umstände der Entstehung der Tetrapoden-Extremität bei. Evolutionstheoretisch scheint nun ein Szenario nahezuliegen, wonach die Beine sich im Wasser entwickelt haben. Extremitäten mit Fingern evolvierten demnach vor dem Landgang. Als Selektionsdruck wird ein besseres Wühlvermögen in vegetationsreichen Uferzonen genannt. Es wurde auch schon die Idee geäußert, mit Beinen sei – wie bei heutigen Amphibien – eine bessere Umklammerung bei der Paarung möglich (Clack 2002, 136). Zur Frage der Selektionsdrücke bei der Entstehung der Tetrapoden wird im dritten Teil noch besonders eingegangen. Es sei aber schon angemerkt, daß man sich hier auf sehr spekulativem Boden befindet. Clack (2002, 136) räumt ein, daß keine der vorgetragenen Hypothesen zum Ursprung der Tetrapodenextremität derzeit testbar sei.
Ungeachtet der Selektionsverhältnisse verkürzt Acanthostega die Kluft zwischen Fischen und auf Land gehenden Tetrapoden. Dies gilt aber nur bei einer groben, undifferenzierten Betrachtung, etwa bei einem bloßen Zusammenzählen von Tetrapodenmerkmalen. Im Detail zeigen sich evolutionstheoretische Probleme, die z. T. schon angesprochen wurden:
- Einzelne Merkmale tauchen markant verändert im Vergleich zu potentiellen Vorläufern auf (z. B. im Bereich des Hirnschädels, oder der Besitz von Fingern).
- Das Handgelenk von Acanthostega war späteren Tetrapoden sehr unähnlich (Clack 2002, 137).
- Neben Merkmalen, in denen Acanthostega abgeleiteter ist als Ichthyostega (Carroll 1992, 60), gibt es auch Eigenschaften, bei denen das Umgekehrte zutrifft.
- Die Wangen und das Schädeldach sind bei Acanthostega und Ichthyostega fest verbunden, im Gegensatz sowohl zu den möglichen Fisch-Vorfahren als auch zu anderen frühen Tetrapoden (Carroll 1992, 60). Das heißt, evolutionär gesehen wäre hier ein Zickzackkurs gefahren worden.
- Carroll (1992, 60) bemerkt: Verschiedene Spezialisierungen schließen die Möglichkeit einer direkten Vorfahrenschaft für spätere Tetrapoden aus. (Eine Zusammenstellung einzigartiger, abgeleiteter und primitiver Merkmale von Acanthostega findet sich bei Coates 1996, 369).
Rückkehr zum Leben im Wasser? Einige Autoren halten es für möglich, daß die ausschließliche Lebensweise im Wasser das Ergebnis eines sekundären Evolutionsweges ist. Acanthostega stamme also von ehemals mehr landlebenden Formen ab. Das schlagkräftigste und schwer zu widerlegende Argument für diese Ansicht sieht Clack im tetrapodenartigen Becken und den großen Oberschenkelknochen von Acanthostega. Es sei schwer vorstellbar, zu welchem Zweck sie sich entwickelt haben sollten, wenn nicht zum Gehen. Ähnliches gelte für die Ausbildung von Fingern (Clack 2002, 135; vgl. Clack & Coates 1995, 366). Außerdem hat Acanthostega die Knochen der Operculogular-Serie am hinteren Ende des Schädels verloren, die im Wasser für Kiemenatmung nützlich sind (wenn auch nicht notwendig, wie andere Fische beweisen). Die Plesiomorphien („Primitivmerkmale“) von Acanthostega (z. B. einfache Wirbelsäule, einfach gebaute Rippen) könnten z. T. auch als sekundäre Wasser-Anpassungen interpretiert werden.
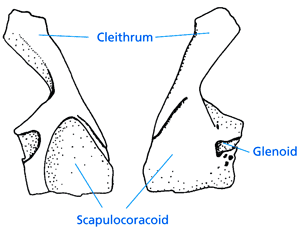 |
| Abb. 16: Innen- (links) und Außenseite des Schultergürtels von Hynerpeton. (Nach Daeschler et al. 1994). |
|
Andererseits sprechen die Anatomie der Arme (ähnlich Eusthenopteron) und des Schwanzes deutlich für eine primäre aquatische Lebensweise; andernfalls müßte man eine gravierende Reversion (Rückentwicklung) annehmen. Kein sekundär wasserbewohnender Vierbeiner hat eine Speiche, die deutlich länger als die Elle wäre; bei der Quastenflosser-Ordnung Osteolepiformes ist das aber die Regel. Als Rückentwicklung müßte auch das ausgeprägte, von Knochen eingeschlossene Sinneskanal-Netzwerk angesehen werden (Coates 1996, 402). Acanthostega besitzt auch Flossenstrahlen, die von der Haut gebildet werden; das macht ein sekundäres Wasserleben unwahrscheinlich (Clack 2002, 128).
Insgesamt entsteht also ein ausgeprägt widersprüchlicher Gesamteindruck. Er wird von Clack (2002, 136) wie folgt auf den Punkt gebracht: „It is at least as difficult to imagine a terrestrial animal with a tail fin like Acanthostega as it is to imagine a primitively aquatic one having evolved a large pelvis and femur.“ Allerdings besteht diese Schwierigkeit nur im Rahmen des evolutionären Paradigmas, denn dort muß nach Selektionsdrücken für die Änderungen des Körperbaus gefragt werden, und genau darin besteht das Problem und das mindert die Plausibilität evolutionärer Szenarien. Der gesamte Merkmalskomplex kann dagegen durchaus als passende Einrichtung für eine bestimmte ökologische Nische in vegetationsreichen Uferzonen verständlich gemacht werden. So kann etwa das starke Becken als Muskelansatzstelle für einen gut ausgebildeten Schwanz dienen, der wiederum ein schnelles Hervorstoßen aus einer Lauerstellung ermöglicht (Clack 2002, 136). Doch eine plausible Ökologie ist noch kein Argument für einen plausiblen Evolutionsvorgang. Wir kommen auf die Problematik der Selektionsdrücke im dritten Teil noch einmal zurück.
|
|
Andere Tetrapoden-Gattungen des Famenniums
Weitere Gattungen des Famenniums sind nur bruchstückhaft bekannt. Ihre Zugehörigkeit zu den Vierbeinern kann nicht als gesichert gelten, da in keinem Fall die vorderen Teile der Extremitäten (Ausbildung von Fingern?) bekannt sind.
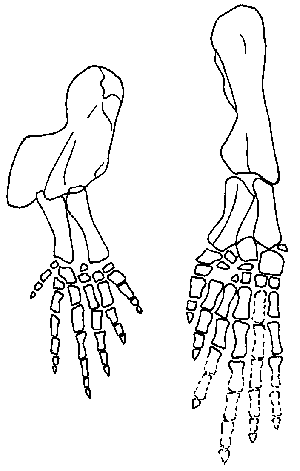 |
| Abb. 17: Rekonstruierte Vorder- (links) und Hinterextremität von Tulerpeton curtum. (Nach Clack 2002). |
|
Von Hynerpeton aus Nordamerika sind Schultergürtel (Abb. 16), das Jugale (ein Wangenknochen des Schädels), ein Teil des Unterkiefers und einige Bauchschuppen fossil überliefert. Daeschler et al. (1994, 640) charakterisieren Hynerpeton als einzigartige Kombination primitiver, abgeleiteter und autapomorpher (spezieller) Merkmale.
Der Schultergürtel besitzt Ansatzstellen für kräftige Muskulatur zur Vor- und Rückwärtsbewegung und Hebung der Vorderextremitäten. Dies gilt als Hinweis dafür, daß diese Gattung sich eher (oder ggf. besser) auf dem Land fortbewegen konnte als andere oberdevonische Vierbeiner (Daeschler et al. 1994, 641). Extremitätenknochen sind allerdings unbekannt, so daß die Fortbewegungsweise nicht bestimmt werden kann (Clack 1997, 230). Vermutlich fehlten innere Kiemen (Daeschler et al. 1994). Insgesamt sind die Überreste zu bruchstückhaft und die Merkmale zu wenig bedeutsam, um klare Schlußfolgerungen über Lebensweise und Evolution zu ziehen.
Tulerpeton stammt aus dem Oberdevon Russlands. Es sind eine rechte Vorder- und Hinterextremität, ein linker Schultergürtel, wenige Teile des Schädels und Bauchschuppen fossil erhalten. Auffallend ist der Besitz von 6 Fingern (Abb. 17). Die relativ schlanken Gliedmaßen ähneln insgesamt deutlich späteren Vierbeinern (Anthracosauriern; „Kohlenechsen“) des Karbons (Carroll 1992, 56; Janvier 1996, 229). Die Knöchelkonstruktion gleicht jedoch insofern Ichthyostega und Acanthostega, als der Fuß weniger Knochen aufweist als spätere Tetrapoden. Schienbein und Wadenbein sind abgeflacht und eher paddelartig, eine Situation, die für heutige Wassersalamander typisch ist (Clack 2002, 129f.). Der Schultergürtel ist robuster als bei Acanthostega und kann daher als weniger „wasserangepaßt“ interpretiert werden. Auch Tulerpeton hatte vermutlich keine inneren Kiemen. Von dieser Gattung ist ebenfalls zu wenig bekannt, um sich ein begründetes Bild von seiner Lebensweise machen zu können.
Von Ventastega aus dem Oberdevon Litauens wurden Schädelteile (Abb. 18) und Teile von Schulter- und Beckengürtel gefunden, die anderen Devon-Tetrapoden wie Acanthostega, Ichthyostega und Tulerpeton weitgehend ähneln (Ahlberg et al. 1994, 303). Die Bezahnung gleicht z. T. eher Panderichthys; Reißzähne auf dem Gaumen werden als fischartig und primitiv eingestuft (Fressen im Wasser) (Clack 2002, 130). Nach Ahlberg et al. (1994, 306) zeigen diverse Schädelmerkmale ohne Zweifel, daß Ventastega einer der primitivsten Vierbeiner ist – dies steht im Kontrast zur stratigraphischen Stellung, da Ventastega ausgerechnet zu den jüngsten Oberdevon-Tetrapoden gehört.
Der Vollständigkeit halber seien aus dem Famennium noch Densignathus, Metaxygnathus und Sinostega erwähnt. Von Densignathus aus Pennsylvania (USA) ist ein Oberkiefer gefunden wurden, der Hynerpeton ähnelt, jedoch etwas kleiner ist (Daeschler 2000, 304).
Metaxygnathus ist biogeographisch bemerkenswert, denn diese Gattung wurde relativ weit entfernt von den anderen oberdevonischen Gattungen in Australien gefunden. Von dieser Gattung ist nur ein Unterkiefer bekannt, dessen Merkmale z. T. tetrapodentypisch sind (Clack 2002, 133).
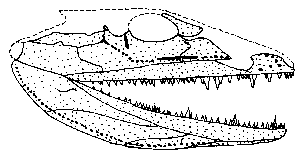 |
| Abb. 18: Rekonstruierter Schädel von Ventastega curonica. (Nach Ahlberg et al. 1994) |
|
Sinostega aus China ist erst im Jahr 2002 beschrieben worden (Zhu et al. 2002) und hat ebenfalls zur Ausweitung des geographischen Bereichs devonischer Tetraopodenfunde beigetragen. Es handelt sich um den ersten Fund aus Asien. Auch von dieser Gattung wurde nur ein Unterkieferbruchstück gefunden, der am ehesten Acanthostega ähnelt. Neuerdings wurde ein tetrapodenähnlicher Oberarmknochen im obersten Oberdevon Pennsylvanias entdeckt (Shubin et al 2004); er ist abgeplattet und ist nicht passend für das Gehen an Land gebaut (vgl. Clack 2004).
Kurze Bewertung der Tetrapoden des Famenniums. Die Tetrapoden des Famenniums sind im postkraniellen Bereich relativ variabel, im Schädelbau jedoch bemerkenswert einheitlich (Ahlberg 1995, 424). Ahlberg sieht dies als typisch für die Stammlinie an vermutet einen Zusammenhang mit einer bestimmten Art des Beuteerwerbs. Die Einheitlichkeit der Schädelform der famennischen Vierbeiner steht im Kontrast zur verlängerten und spezialisierten Schädelform der Elginerpetontidae des (älteren) Frasniums (vgl. 1. Teil in der vorigen Ausgabe).
Ahlberg (1995, 423f.) hält die famennischen Tetrapoden für eine Ansammlung nicht näher verwandter Formen („paraphyletic array“).
|
|
Sauripterus
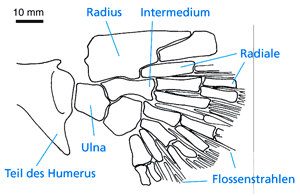 |
| Abb. 19: Flossenskelett von Sauripterus. (Nach Daeschler & Shubin 1997) |
|
Besondere Erwähnung muß noch Sauripterus aus dem Oberdevon, der „Fisch mit Fingern“ finden (Abb. 19). Diese Gattung gehört zur Quastenflosser-Ordnung der Rhizodontida, die phylogenetisch außerhalb einer Gruppe aus Osteolepiformes, Elpistostegalier und Tetrapoden gestellt wird und somit nicht näher mit den Tetrapoden verwandt ist (Daeschler & Shubin 1997). Die Sauripterus-Flosse ist mit 8 fingerartigen radialen Knochen bestückt, die oberflächlich Fingern der frühen Tetrapoden ähneln. Die Fingerknochen sind abgeflacht und relativ steif (Daeschler & Shubin 1997).
Da Sauripterus nicht näher verwandt mit Tetrapoden ist, müssen die fingerartigen Strukturen konvergent (mindestens zweimal unabhängig) entstanden sein; die Brustflosse von Sauripterus kann nicht als Modell für eine Vorstufe von Beinen interpretiert werden (Johanson & Ahlberg 1998, 569). Sauripterus zeigt ebenso wie die Oberdevon-Tetrapoden, daß der Erwerb von Fingern nicht im Zusammenhang mit der Eroberung des Landes gesehen werden muß (Daeschler & Shubin 1997, 133).
|
|
Gesamtbewertung der oberdevonischen Tetrapoden
 |
| Tab 3: Überblick über Änderungen in der Abfolge Eusthenopteron – Panderichthys – Acanthostega (weitgehend nach Clack 2002, 190). Es wird deutlich, daß von Eusthenopteron zu Panderichthys nur relativ geringfügige Änderungen in Richtung Tetrapoden zu verzeichnen sind. Dagegen ist der Sprung von Panderichthys zum Tetrapden Acanthostega beträchtlich. |
|
Insgesamt weisen die oberdevonischen Tetrapoden unterschiedliche Merkmalsmosaike auf, die in phylogenetischen Darstellungen die Annahme vieler Konvergenzen und Reversionen, also zwei- oder mehrfache unabhängige Entstehung und Rückwärtsentwicklungen erfordern. Die Verteilung primitiver und abgeleiteter („höherentwickelter“) Merkmale wechselt von Gattung zu Gattung. Der daher anzunehmende konvergente Ursprung abgeleiteter Merkmale in verschiedenen Linien erschwert die Ermittlung von Verwandtschaftsverhältnissen (Carroll 1995, 390).
Die tetrapodenartigen Formen des Oberdevons tauchen relativ unvermittelt und geographisch weit verbreitet auf (Zhu et al. 2002, 760). Eine graduelle Entwicklung dieser Formen kann bisher nicht belegt werden. Coates et al. (2002, 390) stellen fest, daß es eine große Lücke zwischen Flossen und gefingerten Extremitäten gebe. Ebenso ist die Verbindung zu karbonischen Tetrapoden unklar (s.u.). Angesichts der Formenvielfalt im Oberdevon plädiert Thomson (1993, 48f.) dafür, die frühesten Tetrapoden nicht nur im Oberdevon, sondern im ganzen Devon zu suchen.
Auffällig ist, daß alle oberdevonischen Formen ausnahmslos zusammen mit Fischen abgelagert sind. Dies ist als Indiz dafür zu werten, daß auch die oberdevonischen Tetrapoden wasserlebend waren. Möglicherweise haben diese Formen daher mit der eigentlichen Eroberung des Landes nichts zu tun. Für Janvier (1996, 285) ist klar, daß die Beine ursprünglich nicht für ein Gehen an Land entwickelt wurden, sondern eine spezielle Art von Flossen bildeten, vermutlich für ein Schwimmen in Flußdeltas gegen den Strom oder durch Bereiche mit verottendem Pflanzenmaterial.
Die tetrapodenartigen Merkmale können also möglicherweise durch eine spezialisierte Lebensweise in vegetationsreichen Uferzonen plausibel gemacht werden. Jedenfalls dürfte das letzte Wort über die Lebensweise der oberdevonischen Tetrapoden und tetrapodenähnlichen Formen noch nicht gesprochen sein. Die Entdeckung der Fortbewegungsweise des Quastenflossers Latimeria sollte hier zur Vorsicht mahnen. Entgegen früheren Erwartungen benutzen die heutigen Quastenflosser ihre Extremitäten nicht zum Watscheln (Clack 2002, 102).
|
Die oberdevonischen Tetrapoden
eignen sich nicht als
direkte Vorfahren der karbonischen landlebenden Wirbeltiere. |
|
|
Sind die oberdevonischen Tetrapoden ein „Dead-end“? Vor dem Hintergrund der genannten Daten und Argumente stellt sich die Frage, ob die oberdevonischen wasserlebenden Tetrapoden überhaupt als Vorläufer für die im Karbon überlieferten Land-Tetrapoden geeignet sind. Für die grönländischen Gattungen besteht allgemeine Übereinstimmung in der Einschätzung, daß keine von ihnen direkter Vorfahr späterer landlebender Wirbeltiere ist (Milner et al. 1986, 8; Carroll 1992, 47). Auf diese Frage soll im dritten Teil bei der Besprechung der Karbon-Tetrapoden noch gesondert eingegangen werden, doch können dazu schon an dieser Stelle einige Bemerkungen gemacht werden:
- Ichthyostega scheidet aufgrund vieler spezieller Merkmale als Übergangsform aus.
- Bei Acanthostega kommen alle Merkmale außer den Extremitätenmerkmalen auch bei modernen Fischen vor, die nichts mit dem Landleben zu tun haben (Clack 2002, 137).
- Es ist kein guter Anschluß der oberdevonischen Tetrapoden an unterkarbonische Formen möglich; die meisten Formen des Devons seien wohl – so Clack (2002, 196) – am Ende des Devons ausgestorben. Die karbonischen Tetrapoden gingen auf eine neue Radiation zurück, die zu einer weiten morphologischen und ökologischen Spannbreite geführt habe. (Das wird im dritten Teil behandelt.)
- Alle Oberdevon-Tetrapoden wurden zusammen mit Fischen eingebettet. Das läßt (wie bereits angesprochen) darauf schließen, daß die Tetrapoden des Oberdevons auch wasserlebend waren. Damit ist ihre Deutung als Übergangsformen, die sich auf dem Weg ans Land befinden, geschwächt.
|
Literatur
- Ahlberg PE (1995)
- Elginerpeton pancheni and the Earliest Tetrapod Clade. Nature 373, 420-425
- Ahlberg PE, Luksevics E & Lebedev OA (1994)
- The first tetrapod finds from the Devonian (Upper Famennian) of Latvia. Philos Trans. R. Soc. Lond. B 343, 303-328.
- Ahlberg PE & Milner AR (1994)
- The origin and early diversification of tetrapods. Nature 368, 507-514.
- Carroll RL (1992)
- The primary radiation of terrestrial vertebrates. Annu. Rev. Earth Planet. Sci. 20, 45-84.
- Carroll RL (1993)
- Paläontologie und Evolution der Wirbeltiere. Stuttgart.
- Carroll RL (1995)
- Between fish and amphibian. Nature 373, 389-390.
- Clack JA (1989)
- Discovery of the earliest known tetrapod stapes. Nature 342, 425-430.
- Clack JA (1994a)
- Acanthostega gunnari, a Devonian tetrapod from Greenland; the snout, palate and ventral parts of the braincase, with a discussion of their significance. Meddelelser om Gronland, Geoscience 31, 1-24.
- Clack JA (1994b)
- Earliest known tetrapod braincase and the evolution of the stapes and fenestra ovalis. Nature 369, 392-394.
- Clack JA (1997)
- Devonian tetrapod trackways and trackmakers; a review of the fossils and footprints. Palaeogeogr. Palaeoclim. Palaeoecol. 130, 227-250.
- Clack JA (2001)
- The otoccipital region: origin, ontogeny and the fish-tetrapod transition. In: Ahlberg PE (ed) Major Events in Vertebrate Evolution. London, New York: Taylor & Francis, pp 392-405.
- Clack JA (2002)
- Gaining Ground. The origin and evolution of Tetrapods. Bloomington and Indianapolis: Indiana University Press.
- Clack JA (2003)
- A uniquely specialized ear in a very early tetrapod. Nature 423, 65-69.
- Clack JA (2004)
- From fins to fingers. Science 304, 57-58.
- Clack JA & Coates MI (1995)
- Acanthostega – a primitive aquatic tetrapod? In: Arsenault M, Lelièvre H & Janvier P (eds) 7th International Symposium on Studies on Early Vertebrates. Paris: Bull. Mus. Natl. Hist. Nat., pp 359-372.
- Coates MI (1996)
- The Devonian tetrapod Acanthostega gunnari Jarvik: postcranial anatomy, basal tetrapod interrelationships and patterns of skeletal evolution. Trans. Roy. Soc. Edinburgh 87, 363-421.
- Coates MI & Clack JA (1991)
- Fish-like gills and breathing in the earliest known tetrapod. Nature 352, 234-236.
- Coates MI & Clack JA (1995)
- Romer’s gap: tetrapod origins and terrestriality. In: Arsenault M, Lelièvre H & Janvier P (eds) Studies on early vertebrates (VIIth International Symposium, Miguasha Parc, Quebec). Paris: Bull. Mus. Natl. Hist. Nat., pp 373-388.
- Coates MI, Jeffery JE & Ruta M (2002)
- Fins to limbs: what the fossils say. Evol. Dev. 4, 390-401.
- Daeschler EB & Shubin N (1997)
- Fish with fingers? Nature 391, 133.
- Daeschler EB, Shubin NH, Thomson KS & Amaral WW (1994)
- A Devonian tetrapod from North America. Science 265, 639-642.
- Janvier P (1996)
- Early Vertebrates. Oxford.
- Jarvik E (1996)
- The Devonian tetrapod Ichthyostega. Fossils Strata 40, 1-206.
- Johanson Z & Ahlberg PE (1998)
- A complete primitive rhizodontid from Australia. Nature 394, 569-573.
- Milner AR, Smithson TR, Milner AC, Coates MI & Rolfe WDI (1986)
- The search for early tetrapods. Modern Geology 10, 1-28.
- Shubin NH, Daeschler, EB & Coates MI (2004)
- The early evolution of the tetrapod humerus. Science 304, 90-93.
- Thomson KS (1993)
- The origin of the tetrapods. Am. J. Sci. 293, 33-62.
- Zhu M, Ahlberg PE, Zhao W & Jia L (2002)
- First Devonian tetrapod from Asia. Nature 420, 760-761.
|
|
|  |

